Hepatitis delta virus ribozyme
[1] The ribozyme acts to process the RNA transcripts to unit lengths in a self-cleavage reaction during replication of the hepatitis delta virus, which is thought to propagate by a double rolling circle mechanism.
[2][3] The ribozyme is active in vivo in the absence of any protein factors and was the fastest known naturally occurring self-cleaving RNA at the time of its discovery.
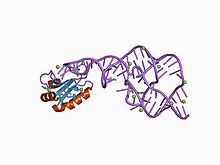
[4] The crystal structure of this ribozyme has been solved using X-ray crystallography and shows five helical segments connected by a double pseudoknot.
[7][8][6][9][10][11] The grouping is probably a result of convergent evolution: Deltavirus found outside of humans also possess a DV ribozyme, and no horizontal gene transfer scenarios proposed can yet explain this.
[1] However, although it is well established that the N3 of C75 has a pKa perturbed from its normal value of 4.45 and is closer to about 6.15 or 6.40,[16][17] it is not neutral enough to act as a general base catalyst.
However, it is likely that the structural environment within the ribozyme, which includes a desolvated active site cleft, provides negative electrostatic potential that could perturb the pKa of cytosine enough to act as a Lewis acid.
[20][21][22][23] In addition to Lewis acid stabilization of the 5′-hydroxyl leaving group, it is also now accepted that the HDV ribozyme can use a metal ion to assist in activation of the 2′-hydroxyl for attack on the U(-1) nucleotide.
As limited by the rapid self-cleaving nature of HDV ribozyme, the previous ribonuclease experiments were performed on the 3′ product of self-cleavage rather than the precursor.
[26] The flanking sequence sequesters the ribozyme in a kinetic trap during transcription and results in the extremely diminished self-cleavage rate.
[26] Second, a series of DNA oligomer complementary to different regions of AS1/2 were used to rescue the ribozyme activity; the results confirms the inhibitory roles of AS1/2.
The ribozyme sequence will efficiently cleave itself with no downstream requirements, as the -1 nucleotide is invariant, leaving a 2′–3′ cyclic phosphate that can easily be removed by treatment with a phosphatase or T4 polynucleotide kinase.