Jurassic
During a tour of the region in 1795, German naturalist Alexander von Humboldt recognized carbonate deposits within the Jura Mountains as geologically distinct from the Triassic aged Muschelkalk of southern Germany, but he erroneously concluded that they were older.
In this book, Brongniart used the phrase terrains jurassiques when correlating the "Jura-Kalkstein" of Humboldt with similarly aged oolitic limestones in Britain, thus coining and publishing the term "Jurassic".
The French palaeontologist Alcide d'Orbigny in papers between 1842 and 1852 divided the Jurassic into ten stages based on ammonite and other fossil assemblages in England and France, of which seven are still used, but none has retained its original definition.
The German palaeontologist Albert Oppel in his studies between 1856 and 1858 altered d'Orbigny's original scheme and further subdivided the stages into biostratigraphic zones, based primarily on ammonites.
[3][8] Albert Oppel in 1858 named the Pliensbachian Stage after the hamlet of Pliensbach in the community of Zell unter Aichelberg in the Swabian Alb, near Stuttgart, Germany.
The GSSP for the base of the Pliensbachian is found at the Wine Haven locality in Robin Hood's Bay, Yorkshire, England, in the Redcar Mudstone Formation, and was ratified in 2005.
[12] The Bathonian is named after the city of Bath, England, introduced by Belgian geologist d'Omalius d'Halloy in 1843, after an incomplete section of oolitic limestones in several quarries in the region.
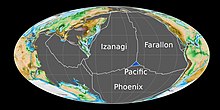
[26] Beginning in the Early Jurassic, the Boreal Ocean was connected to the proto-Atlantic by the "Viking corridor" or Transcontinental Laurasian Seaway, a passage between the Baltic Shield and Greenland several hundred kilometers wide.
[33] Madagascar and Antarctica began to rift away from Africa during the late Early Jurassic in association with the eruption of the Karoo-Ferrar large igneous provinces, opening the western Indian Ocean and beginning the fragmentation of Gondwana.
[62] The Pangaean interior had less severe seasonal swings than in previous warm periods as the expansion of the Central Atlantic and Western Indian Ocean provided new sources of moisture.
[67] Groups affected include ammonites,[68] ostracods,[65][69] foraminifera,[70][71] bivalves,[65] cnidarians, and especially brachiopods,[72][73][74] for which the TOAE represented one of the most severe extinctions in their evolutionary history.
[86] Dicroidium, a corystosperm seed fern that was a dominant part of Gondwanan floral communities during the Triassic, also declined at the Triassic–Jurassic boundary, surviving as a relict in Antarctica into the Early Jurassic.
[96][97] The oldest unambiguous record of Pinaceae is the pine cone Eathiestrobus, known from the Late Jurassic (Kimmeridgian) of Scotland, which remains the only known unequivocal fossil of the group before the Cretaceous.
[99][89] The earliest record of the yew family (Taxaceae) is Palaeotaxus rediviva, from the Hettangian of Sweden, suggested to be closely related to the living Austrotaxus, while Marskea jurassica from the Middle Jurassic of Yorkshire, England and material from the Callovian–Oxfordian Daohugou Bed in China are thought to be closely related to Amentotaxus, with the latter material assigned to the modern genus, indicating that Taxaceae had substantially diversified by the end of the Jurassic.
[103][104] Umaltolepis from the Jurassic of Asia has strap-shaped ginkgo-like leaves with highly distinct reproductive structures with similarities to those of peltasperm and corystosperm seed ferns, has been suggested to be a member of Ginkgoales sensu lato.
The Williamsoniaceae are thought to have had a divaricate branching habit, similar to that of living Banksia, and adapted to growing in open habitats with poor soil nutrient conditions.
Several groups of insects that bear long proboscis, including extinct families such as kalligrammatid lacewings[109] and extant ones such as acrocerid flies,[110] are suggested to have been pollinators of bennettitales, feeding on nectar produced by bennettitalean cones.
[133] The moss Kulindobryum from the Middle Jurassic of Russia, which was found associated with dinosaur bones, is thought to be related to the Splachnaceae, which grow on animal caracasses.
An unnamed protosuchid known from teeth from the Early Jurassic of Arizona represents the earliest known herbivorous crocodylomorph, an adaptation that appeared several times during the Mesozoic.
[168] Within Thalattosuchia, the Metriorhynchidae became highly adapted for life in the open ocean, including the transformation of limbs into flippers, the development of a tail fluke, and smooth, scaleless skin.
[171] Razanandrongobe from the Middle Jurassic of Madagascar has been suggested to represent the oldest record of Notosuchia, a primarily Gondwanan clade of mostly terrestrial crocodylomorphs, otherwise known from the Cretaceous and Cenozoic.
[179] Rhynchocephalians reached their highest morphological diversity in their evolutionary history during the Jurassic, occupying a wide range of lifestyles, including the aquatic pleurosaurs with long snake-like bodies and reduced limbs, the specialized herbivorous eilenodontines, as well as the sapheosaurs which had broad tooth plates indicative of durophagy.
[206] While the earliest salamander-line amphibians are known from the Triassic,[207] crown group salamanders first appear during the Middle to Late Jurassic in Eurasia, alongside stem-group relatives.
[208] Early representatives of crown group salamanders include Chunerpeton, Pangerpeton and Linglongtriton from the Middle to Late Jurassic Yanliao Biota of China.
[213] The fourth group of lissamphibians, the extinct salamander-like albanerpetontids, first appeared in the Middle Jurassic, represented by Anoualerpeton priscus from the Bathonian of Britain, as well as indeterminate remains from equivalently aged sediments in France and the Anoual Formation of Morocco.
Archaic "palaeoniscoid" fish, which were common in both marine and freshwater habitats during the preceding Triassic declined during the Jurassic, being largely replaced by more derived actinopterygian lineages.
[249] The oldest record of angelsharks (Squatiniformes) is Pseudorhina from the Late Jurassic (Oxfordian–Tithonian) of Europe, which already has a bodyform similar to members of the only living genus of the order, Squatina.
[250] The oldest known remains of Carcharhiniformes, the largest order of living sharks, first appear in the late Middle Jurassic (Bathonian) of the western Tethys (England and Morocco).
[287] Echinoids (sea urchins) underwent substantial diversification beginning in the Early Jurassic, primarily driven by the radiation of irregular (asymmetrical) forms, which were adapting to deposit feeding.
[294] Jurassic barnacles were of low diversity compared to present,[295] but several important evolutionary innovations are known, including the first appearances of calcite shelled forms and species with an epiplanktonic mode of life.