Lambda phage
Enterobacteria phage λ (lambda phage, coliphage λ, officially Escherichia virus Lambda) is a bacterial virus, or bacteriophage, that infects the bacterial species Escherichia coli (E. coli).
[2] The wild type of this virus has a temperate life cycle that allows it to either reside within the genome of its host through lysogeny or enter into a lytic phase, during which it kills and lyses the cell to produce offspring.
Usually, a "lytic cycle" ensues, where the lambda DNA is replicated and new phage particles are produced within the cell.
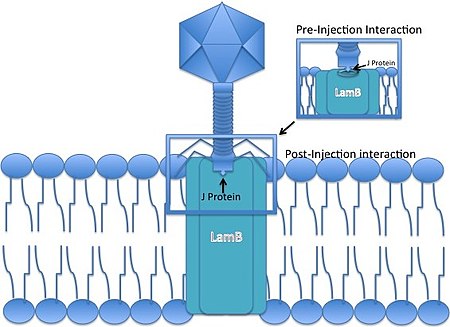
The tail of lambda phages is made of at least 6 proteins (H, J, U, V, Stf, Tfa) and requires 7 more for assembly (I, K, L, M, Z, G/T).
Low temperature, starvation of the cells and high multiplicity of infection (MOI) are known to favor lysogeny (see later discussion).
This is the lifecycle that the phage follows following most infections, where the cII protein does not reach a high enough concentration due to degradation, so does not activate its promoters.
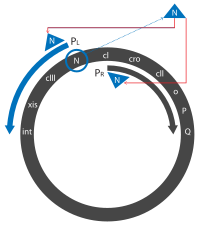
O and P are responsible for initiating replication, and Q is another antiterminator that allows the expression of head, tail, and lysis genes from PR’.
Gam is also important in that it inhibits the host RecBCD nuclease from degrading the 3’ ends in rolling circle replication.
[18] The lysogenic lifecycle begins once the cI protein reaches a high enough concentration to activate its promoters, after a small number of infections.
In response to stress, the activated prophage is excised from the DNA of the host cell by one of the newly expressed gene products and enters its lytic pathway.
The sequence of the bacterial att site is called attB, between the gal and bio operons, and consists of the parts B-O-B', whereas the complementary sequence in the circular phage genome is called attP and consists of the parts P-O-P'.
Both Int and IHF bind to attP and form an intasome, a DNA-protein-complex designed for site-specific recombination of the phage and host DNA.
Any situation where a lysogen undergoes DNA damage or the SOS response of the host is otherwise stimulated leads to induction.
[22] Prophage reactivation in phage λ appears to occur by a recombinational repair process similar to that of MR.
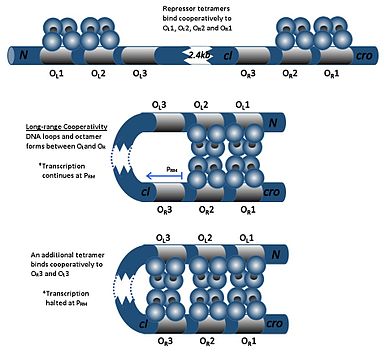
The repressor found in the phage lambda is a notable example of the level of control possible over gene expression by a very simple system.
This autonegative regulation ensures a stable minimum concentration of the repressor molecule and, should SOS signals arise, allows for more efficient prophage induction.
The latter is determined solely by the activation of RecA in the SOS response of the cell, as detailed in the section on induction.
The former will also be affected by this; a cell undergoing an SOS response will always be lysed, as no cI protein will be allowed to build up.
cIII appears to stabilize cII, both directly and by acting as a competitive inhibitor to the relevant proteases.
Computer modeling and simulation suggest that random processes during infection drive the selection of lysis or lysogeny within individual cells.
[28] Some of its uses include its application as a vector for the cloning of recombinant DNA; the use of its site-specific recombinase (int) for the shuffling of cloned DNAs by the gateway method;[29] and the application of its Red operon, including the proteins Red alpha (also called 'exo'), beta and gamma in the DNA engineering method called recombineering.
Lambda phage will enter bacteria more easily than plasmids, making it a useful vector that can either destroy or become part of the host's DNA.
[31] Lambda phage can also be manipulated and used as an anti-cancer vaccine that targets human aspartyl (asparaginyl) β-hydroxylase (ASPH, HAAH), which has been shown to be beneficial in cases of hepatocellular carcinoma in mice.