Neural Darwinism
Neural Darwinism is a biological, and more specifically Darwinian and selectionist, approach to understanding global brain function, originally proposed by American biologist, researcher and Nobel-Prize recipient[1] Gerald Maurice Edelman (July 1, 1929 – May 17, 2014).
[4] The development of neural Darwinism was deeply influenced by work in the fields of immunology, embryology, and neuroscience, as well as Edelman's methodological commitment to the idea of selection as the unifying foundation of the biological sciences.
In accordance with principles of population biology and Darwin's theory of natural selection – as opposed to the top-down algorithmic and computational approaches that dominated a nascent cognitive psychology at the time.
Edelman was a medical researcher, physical chemist, immunologist, and aspiring neuroscientist when he was awarded the 1972 Nobel Prize in Physiology or Medicine (shared with Rodney Porter of Great Britain).
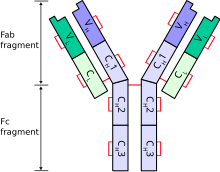
Edelman would draw inspiration from the mechano-chemical aspects of antigen/antibody/lymphocyte interaction in relation to recognition of self-nonself; the degenerate population of lymphocytes in their physiological context; and the bio-theoretical foundations of this work in Darwinian terms.
By 1974, Edelman felt that immunology was firmly established on solid theoretical grounds descriptively, was ready for quantitative experimentation, and could be an ideal model for exploring evolutionary selection processes within an observable time period.
He proceeds to demonstrate how the "noise" of the computational and algorithmic approach is actually beneficial to a somatic selective system by providing a wide, and degenerate, array of potential recognition elements.
This property of degeneracy makes the system more adaptively robust in the face of unforeseen contingencies: When one particular solution fails unexpectedly, there are other unaffected pathways that can be engaged in pursuit of the same end.
Early on, Edelman spends considerable time contrasting degeneracy vs. redundancy, bottom-up vs. top-down processes, and selectionist vs. instructionist explanations of biological phenomena.
Edelman's commitment to the Darwinian underpinnings of biology, his emerging understanding of the evolutionary relationships between the two molecules he had worked with, and his background in immunology lead him to become increasingly critical and dissatisfied with attempts to describe the operation of the nervous system and brain in computational or algorithmic terms.
Edelman acknowledges that there is a conservation of phylogenetic organization and structure within the vertebrate nervous system, but also points out that locally natural diversity, variation and degeneracy abound.
The modern synthesis was greatly accelerated and expanded with the rise of the genomic sciences, molecular biology, as well as, advances in computational techniques and the power to model population dynamics.
[22] Edelman, like Lewontin, seeks a complete description of the transformations (T) that take us from:[23] Lewontin's exploration of these transformations between genomic and phenotypic spaces was in terms of key selection pressures that sculpt the organism over geological evolutionary time scales; but, Edelmans approach is more mechanical, and in the here and now – focusing on the genetically constrained mechano-chemistry of the selection processes that guide epigenetic behaviors on the part of cells within the embryo and adult over developmental time.
The expression of the CAMs and SAMs is under genetic control, but the distribution of these molecules on the cell membrane and extracellular matrix is historically contingent upon epigenetic events, serving as one of the primary bases for generating pre-existing diversity within the nervous system and other tissues.
Edelman outright rejects the notion of a homunculus, describing it as a "close cousin of the developmental electrician and the neural decoder", artifacts of the observer-centralized top-down design logic of information processing approaches.
Edelman properly points out that "it is probably a safe guess that most neurobiologists would view the homunculus as well as dualist solutions (Popper and Eccles 1981) to the problems of subjective report as being beyond scientific consideration.
The third part attempts to build a temporally coherent model of globally unitary cognitive function and behavior that emerges from the bottom up through the interactions of the neuronal groups in real-time.
Edelman organized key ideas of the TNGS theory into three main tenets: The primary repertoire is formed during the period from the beginning of neurulation to the end of apoptosis.
The secondary repertoire extends over the period synaptogenesis and myelination, but will continue to demonstrate developmental plasticity throughout life, albeit in a diminished fashion compared to early development.
The two repertoires deal with the issue of the relationship between genetic and epigenetic processes in determining the overall architecture of the neuroanatomy – seeking to reconcile nature, nurture, and variability in the forming the final phenotype of any individual nervous system.
There is no point-to-point wiring that carries a neural code through a computational logic circuit that delivers the result to the brain because Variation is the inevitable outcome of developmental dynamics.
Reentrant signalling is an attempt to explain how "coherent temporal correlations of the responses of sensory receptor sheets, motor ensembles, and interacting neuronal groups in different brain regions occur".
It seeks to provide an explanation of how the diversity of neuronal group phenotypes emerge from the organism's genotype via genetic and epigenetic influences that manifest themselves mechano-chemically at the cell surface and determine connectivity.
Edelman's notion of the secondary repertoire heavily borrows from work of Jean-Pierre Changeux, and his associates Philippe Courrège and Antoine Danchin – and, their theory of selective stabilization of synapses.
Edelman called it "reentry" and proposes a model of reentrant signaling whereby a disjunctive, multimodal sampling of the same stimulus event correlated in time that make possible sustained physiological entrainment of distributed neuronal groups into temporally stable global behavioral units of action or perception.
Simultaneously, each sensory modality is also being sent to the cortex in parallel, for higher-order reflective analysis, multimodal sensorimotor association, and the engagement of the slow modulatory response that will fine-tune the subcortical reflexes.
"Neither the original theory nor simulated recognition automata deal in satisfactory detail with the successive ordering of events in time mediated by the several major brain components that contribute to memory, particularly as it relates to consciousness.
[45] It has been suggested by Chase Herrmann-Pillath that Friedrich Hayek had earlier proposed a similar idea in his book The Sensory Order: An Inquiry into the Foundations of Theoretical Psychology, published in 1952.
A recent review by Fernando, Szathmary and Husbands explains why Edelman's neural Darwinism is not Darwinian because it does not contain units of evolution as defined by John Maynard Smith.
[48] A recent theory called evolutionary neurodynamics being developed by Eors Szathmary and Chrisantha Fernando has proposed several means by which true replication may take place in the brain.