Site-specific recombinase technology
In the late 1980s gene targeting in murine embryonic stem cells (ESCs) enabled the transmission of mutations into the mouse germ line, and emerged as a novel option to study the genetic basis of regulatory networks as they exist in the genome.
Although meganucleases, zinc-finger- and transcription activator-like effector nucleases (ZFNs and TALENs) are actual tools supporting HR, it was the availability of site-specific recombinases (SSRs) which triggered the rational construction of cell lines with predictable properties.
[5] The founding member of the YR family is the lambda integrase, encoded by bacteriophage λ, enabling the integration of phage DNA into the bacterial genome.
Early members of the SR family are closely related resolvase / DNA invertases from the bacterial transposons Tn3 and γδ, which rely on a catalytic serine responsible for attacking the scissile phosphate to form a 5'-phosphoserine linkage.
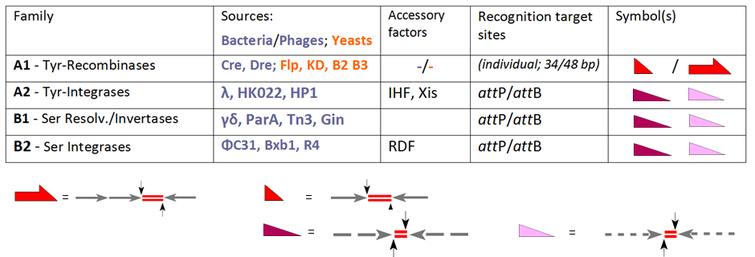
Nowadays the standard activity of each enzyme determines its classification reserving the general term "recombinase" for family members which, per se, comprise all three routes, INT, RES and INV: Our table extends the selection of the conventional SSR systems and groups these according to their performance.
These features account for relevant differences: In order to streamline this chapter the following implementations will be focused on two recombinases (Flp and Cre) and just one integrase (PhiC31) since their spectrum covers the tools which, at present, are mostly used for directed genome modifications.
[10] Relative to Flp-RMCE, which requires two different ("heterospecific") FRT-spacer mutants, the reaction partner (attB) of the first reacting attP site is hit arbitrarily, such that there is no control over the direction the donor cassette enters the target (cf.
One successful strategy for inducing specific temporal Cre activity involves fusing the enzyme with a mutated ligand-binding domain for the human estrogen receptor (ERt).
Recent extensions of these general concepts led to generating the "Cre-zoo", i.e. collections of hundreds of mouse strains for which defined genes can be deleted by targeted Cre expression.
[3] In its natural host (S. cerevisiae) the Flp/FRT system enables replication of a "2μ plasmid" by the inversion of a segment that is flanked by two identical, but oppositely oriented FRT sites ("flippase" activity).
This inversion changes the relative orientation of replication forks within the plasmid enabling "rolling circle"—amplification of the circular 2μ entity before the multimeric intermediates are resolved to release multiple monomeric products.
[6] An additional advantage is the fact, that simple rules can be applied to generate heterospecific FRT sites which undergo crossovers with equal partners but nor with wild type FRTs.
The traditional, laborious "tag-and-exchange" procedures relied on two successive homologous recombination (HR-)steps, the first one ("HR1") to introduce a tag consisting of a selection marker gene.
[6] Further restrictions are imposed by the fact that irreversibility does not permit standard multiplexing-RMCE setups including "serial RMCE" reactions, i.e., repeated cassette exchanges at a given genomic locus.
As one of the most important platforms for dealing with vertebrate gene functions on a large scale, genome-wide genetic resources of mutant murine ES cells have been established.
To this end four international programs aimed at saturation mutagenesis of the mouse genome have been founded in Europe and North America (EUCOMM, KOMP, NorCOMM, and TIGM).
[14] The relevant technologies have now reached a level permitting their extension to other mammalian species and to human stem cells, most prominently those with an iPS (induced pluripotent) status.