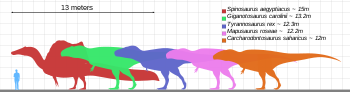
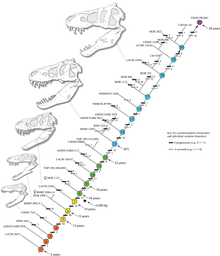
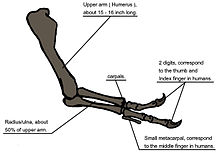
Tyrannosaurus
Fossils are found in a variety of rock formations dating to the latest Campanian-Maastrichtian ages of the late Cretaceous period, 72.7 to 66 million years ago, with isolated specimens possibly indicating an earlier origin in the middle Campanian.
Cope believed the fragments belonged to an "agathaumid" (ceratopsid) dinosaur, and named them Manospondylus gigas, meaning "giant porous vertebra", in reference to the numerous openings for blood vessels he found in the bone.
Originally thought to belong to a hadrosaurid, examination of the footprint revealed a large 'heel' unknown in ornithopod dinosaur tracks, and traces of what may have been a hallux, the dewclaw-like fourth digit of the tyrannosaur foot.
[23][24] A set of footprints in Glenrock, Wyoming dating to the Maastrichtian stage of the Late Cretaceous and hailing from the Lance Formation were described by Scott Persons, Phil Currie and colleagues in 2016, and are believed to belong to either a juvenile T. rex or the dubious tyrannosaurid Nanotyrannus lancensis.
[51][52] The premaxillary teeth, four per side at the front of the upper jaw, were closely packed, D-shaped in cross-section, had reinforcing ridges on the rear surface, were incisiform (their tips were chisel-like blades) and curved backwards.
Various functions have been proposed for these foramina, such as a crocodile-like sensory system[55] or evidence of extra-oral structures such as scales or potentially lips,[56][57][58] with subsequent research on theropod tooth wear patterns supporting such a proposition.
This isolated right lacrimal was originally collected alongside the holotype specimen of Deinosuchus rugosus, a giant crocodylian, and remained undescribed until its re-identification as belonging to a tyrannosaurid theropod in the 1980s by paleontologist Dale Russell.
Differences of general body proportions representing robust and gracile morphotypes were also used as a line of evidence, in addition to the number of small, slender incisiform teeth in the dentary, as based on tooth sockets.
[101][102][103] Peter Larson continued to support the hypothesis that N. lancensis was a separate but closely related species, based on skull features such as two more teeth in both jaws than T. rex; as well as proportionately larger hands with phalanges on the third metacarpal and different wishbone anatomy in an undescribed specimen.
[111] The same year, Carr published a paper on T. rex's growth history, finding that CMNH 7541 fit within the expected ontogenetic variation of the taxon and displayed juvenile characteristics found in other specimens.
[126] The subsequent discovery of the giant species Yutyrannus huali, also from the Yixian, showed that even some large tyrannosauroids had feathers covering much of their bodies, casting doubt on the hypothesis that they were a size-related feature.
[127] A 2017 study reviewed known skin impressions of tyrannosaurids, including those of a Tyrannosaurus specimen nicknamed "Wyrex" (HMNS 2006.1743.01, formerly known as BHI 6230) which preserves patches of mosaic scales on the tail, hip, and neck.
[135] In 1915, convinced that the creature stood upright, Henry Fairfield Osborn, former president of the American Museum of Natural History, further reinforced the notion in unveiling the first complete T. rex skeleton arranged this way.
[136] By 1970, scientists realized this pose was incorrect and could not have been maintained by a living animal, as it would have resulted in the dislocation or weakening of several joints, including the hips and the articulation between the head and the spinal column.
The heavy build of the arm bones, strength of the muscles, and limited range of motion may indicate a system evolved to hold fast despite the stresses of a struggling prey animal.
[153] Barrick and Showers have defended their conclusions in subsequent papers, finding similar results in another theropod dinosaur from a different continent and tens of millions of years earlier in time (Giganotosaurus).
[160] In the March 2005 issue of Science, Mary Higby Schweitzer of North Carolina State University and colleagues announced the recovery of soft tissue from the marrow cavity of a fossilized leg bone from a T. rex.
Paleontologist Hans Larsson of McGill University in Montreal, who was not part of the studies, called the finds "a milestone", and suggested that dinosaurs could "enter the field of molecular biology and really slingshot paleontology into the modern world".
[172] A 2002 report used a mathematical model (validated by applying it to three living animals: alligators, chickens, and humans; and eight more species, including emus and ostriches[172]) to gauge the leg muscle mass needed for fast running (over 40 km/h or 25 mph).
[172] Proposed top speeds exceeded 40 kilometers per hour (25 mph) for Tyrannosaurus, but were deemed infeasible because they would require exceptional leg muscles of approximately 40–86% of total body mass.
Compared to more basal groups of theropods in the study, tyrannosaurs like Tyrannosaurus itself showed a marked increase in foraging efficiency due to reduced energy expenditures during hunting or scavenging.
Additionally, the research, in conjunction with studies that show tyrannosaurs were more agile than other large-bodied theropods, indicates they were quite well-adapted to a long-distance stalking approach followed by a quick burst of speed to go for the kill.
[181] Rare fossil footprints and trackways found in New Mexico and Wyoming that are assigned to the ichnogenus Tyrannosauripus have been attributed to being made by Tyrannosaurus, based on the stratigraphic age of the rocks they are preserved in.
[199][200] According to scientists assessing the Dino Gangs program, the evidence for pack hunting in Tarbosaurus and Albertosaurus is weak and based on group skeletal remains for which alternate explanations may apply (such as drought or a flood forcing dinosaurs to die together in one place).
[203] Pathologies of other Tyrannosaurus specimens have been suggested as evidence of conspecific attack, including "Wyrex" with a hole penetrating its jugual and severe trauma on its tail that shows signs of bone remodeling (not regrowth).
[241] A subsequent study found that while trichomoniasis has many attributes of the model proposed (osteolytic, intra oral) several features make the assumption that it was the cause of death less supportable by evidence.
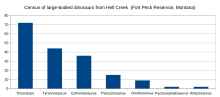
Tyrannosaurus shared this ecosystem with ceratopsians Leptoceratops, Torosaurus, and Triceratops, the hadrosaurid Edmontosaurus annectens, the parksosaurid Thescelosaurus, the ankylosaurs Ankylosaurus and Denversaurus, the pachycephalosaurs Pachycephalosaurus and Sphaerotholus, and the theropods Ornithomimus, Struthiomimus, Acheroraptor, Dakotaraptor, Pectinodon and Anzu.
[246] In its southern range, specifically based on remains discovered from the North Horn Formation of Utah, Tyrannosaurus rex lived alongside the titanosaur Alamosaurus, the ceratopsid Torosaurus and the indeterminate troodontids and hadrosaurids.
Tyrannosaurus have also been discovered from the Javelina Formation of Texas,[68] where the remains of the titanosaur Alamosaurus, the ceratopsid Bravoceratops, the pterosaurs Quetzalcoatlus and Wellnhopterus, and possible species of troodontids and hadrosaurids are found.
[257] The authors of the original publication replied that while they agree that their reported uncertainties were probably too small, their framework is flexible enough to accommodate uncerainty in physiology, and that their calculations do not depend on short-term changes in population density and geographic range, but rather on their long-term averages.