[14] Further gene duplication events of this ancestral TIM barrel led to diverging enzymes possessing the functional diversity observed today.
[3] TIM barrels contain 200-250 amino acid residues,[2] folded into 8 alpha helices (α-helices) and 8 beta strand (β-strands).
Short loops typically connect the α and β secondary structures, forming a (βα)8 repeat topology.
TIM barrels contain two distinct buried regions, where amino acid residues are completely enveloped by their neighbors and lack access to solvent.
Due to the pleated nature of β-strands, alternate residues along a strand are almost evenly split between the pore (53%) and core (47%).
Partially-folded βαβα modules, called foldons, would be energetically stabilized by polar pore residues during this stage of folding.
In this case however, the authors credited branched aliphatic amino acids (valine, leucine, and isoleucine) for foldon stability.
These clamps (or hydrophobic side chain bridge analogs) are conserved in 3 indole-3-glycerolphosphate synthase TIM barrel orthologs from the bacterial and archaeal kingdoms, implying they arose in their last common ancestor and have been preserved for over a billion years.
Bacillus subtilis Orotidine 5'-phosphate decarboxylase (PDB: 1DBT) is a TIM barrel protein displaying 4 α-helices in place of the βα loops typically present at the C-terminal of the β-barrel (residues 35-42, 89-91, 126-133, and 215-219).
Lactococcus lactis Dihydroorotate dehydrogenase A (DHODA) (PDB: 2DOR) is an example of a TIM barrel possessing β-sheets and extended loops over the C-terminal end of the β-barrel.
DHODA catalyzes the oxidation of dihydroorotate to orotate, which is part of the de novo uridine 5'-monophosphate (UMP) synthesis pathway.
Chemical denaturation of several natural[27][28] and 2 designed TIM barrel variants[28] invariably involves a highly populated equilibrium intermediate.
The kinetic intermediates that appear after dilution from highly denaturing solutions involve an early misfolded species that must at least partially unfold to access the productive folding pathway.
[29] Native-centric molecular dynamics simulations recapitulate the experimental results and point the way to testable computational models for complex folding mechanisms.
[30] TIM barrel proteins possess an unusually high sequence plasticity, forming large families of orthologous and paralogous enzymes in widely divergent organisms.
This plasticity suggests a sequence landscape that allows for protein adaptation to a variety of environmental conditions, largely independent of phylogenetic history, while maintaining function.
Of the approximately 200 residues required to fully form a TIM barrel, about 160 are considered structurally equivalent between different proteins sharing this fold.
Recently, it has been demonstrated that catalytic loops can be exchanged between different TIM barrel enzymes as semiautonomous units of functional groups.
In the early 1990s, it was noted that all TIM barrel structures solved at the time were enzymes, indicating divergence from a common ancestor.
suggested that A common phosphate binding site, formed by a small α-helix and TIM barrel loops-7/8, strongly indicated divergent evolution.
A good example of 2-fold internal symmetry is observed in the enzymes ProFAR isomerase (HisA) and imidazole glycerol phosphate synthase (HisF) of the Thermotoga maritima histidine biosynthesis pathway.
More interestingly, the loops on the C terminal ends of both HisA and HisF showed a twofold repeated pattern, suggesting that their common ancestor also possessed 2-fold internal symmetry.
[15][16] The crystal structure of HisF-C***C revealed a 2-fold symmetric TIM barrel, validating the possibility of natural domain fusion.
[17] These experiments led to the proposal of a novel means of diversification and evolution of TIM-barrel enzymes through the exchange of (βα)4 half-barrel domains amongst preexisting TIM barrels.
Although they displayed circular dichroism spectra consistent with αβ proteins and some cooperative folding characteristics, all Octarellin series peptides were insoluble, and had to be resolubilized from inclusion bodies for further characterization.
Despite these advances, all proteins in this family displayed molten characteristics when analyzed using NMR (nuclear magnetic resonance), and further work to solve their structures could not be pursued.
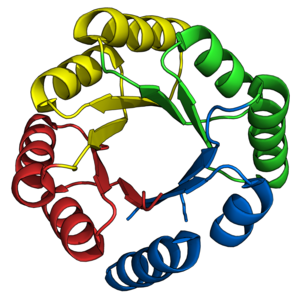
Triose-phosphate isomerase (TIM)
isolated from chicken muscles (
PDB
:
1TIM
), the archetypal TIM barrel enzyme.
(A)
Cartoon representation of the TIM barrel structure. α-helices are colored teal, β-strands are colored orange, and loops are colored green. Note that the C-terminal ends of β-strands are depicted with arrowheads.
(B)
Core and pore regions are highlighted. Amino acid residues belonging to the pore are colored blue. Amino acid residues belonging to the core are colored orange. Note that the TIM barrel is depicted in a
top-down
view, where the C-terminal ends of the β-barrel are pointed towards the reader.
TIM barrel topology.
α-helices are colored teal, loops are colored green, and β-strands are colored in two shades of orange. Lighter shades indicate residues pointing
inward
, towards the barrel pore. Darker shades indicate residues pointing
outward
, towards the barrel core. Cyan lines depict an example backbone β-barrel hydrogen bonding network. Note that side-chain hydrogen bonding networks are not depicted here. Interior β-barrel residues (pore residues) display a 4-fold geometric symmetry, despite emerging from an 8-strand β-barrel. This symmetry is illustrated as two example "layers" in red and blue. Each layer contains 4 residues that point towards the pore, and lie on the same plane perpendicular to the barrel axis. The shear number for TIM barrels is always 8, and is illustrated in magenta. Some TIM barrels naturally adopt, or are designed to adopt, two or four-fold symmetry. Example asymmetric units are also highlighted. This figure has been adapted with permission from previously published work.
[
23
]
Example salt bridge network
in 2-deoxyribose-5-phosphate aldolase (
PDB
:
1P1X
). Interactions are shown as cyan dashed lines. Polar residues are colored green. Polar amino acids aspartate (D), glutamate (E), lysine (K), and arginine (R), are shown here.
Examples of structural inserts at TIM barrel loop and N/C-terminal regions.
(A)
The
Bacillus subtilis
Orotidine 5′-monophosphate decarboxylase (
PDB
:
1DBT
). Orotidine 5'-monophosphate is colored green. α-helical inserts are colored teal. The catalytic arginine residue (R215) is displayed as sticks.
(B)
Mycobacterium tuberculosis
bifunctional histidine/tryptophan biosynthesis isomerase (PriA) (
PDB
:
2Y85
). CdRP, the product of the TrpF reaction, is colored green. β-strand/loop interchangeable structures are colored orange.
(C)
Lactococcus lactis
dihydroorotate dehydrogenase A (DHODA) (
PDB
:
2DOR
). β-strands forming a sheet are colored orange. Extended loops are colored green. The cavity formed by these structures is displayed as a blue mesh. The product orotate is colored magenta. the cofactor FMN is colored pink.
(D)
Methylophilus methylotrophus
trimethylamine dehydrogenase (
PDB
:
2TMD
).
[
26
]
The Rossmann fold domain is colored according to secondary structural elements. Cofactor FMN is colored magenta. The [4Fe-4S]
+
is colored red. Note that substrate/product were not crystallized.
The reaction coordinate diagram for SsIGPS at pH 7.8 and 25°C. The refolding reaction begins in the unfolded,
U
state, initially misfolds to the
I
BP
intermediate state, partially unfolds to reach the
I
A
intermediate state whose conversion to the subsequent
I
B
intermediate state is rate-limiting. The final step is the conversion of
I
B
to the native state, N. The
I
A
and
I
B
kinetic intermediates correspond to the intermediate observed in equilibrium unfolding studies. The ordinate represents the free energy of each state in the folding reaction mechanism in kcal mol
−1
. The abscissa represents the dependence of the difference in free energy between 2 states on the denaturant concentration and is proportional to the change in buried surface, referenced to the
U
state. The kinetic folding mechanism, illustrating the flow of the unfolded protein to the native conformation is shown beneath the reaction coordinate diagram.
Experimentally derived fitness landscapes mapped from point mutations represent single steps from WT sequence. Despite significant divergence of WT in sequence space, the fitness landscapes of IGPS orthologues remain correlated (dashed lines). Rather than traditional two-dimensional heatmaps, fitness values are displayed on a three-dimensional pinwheel, highlighting the wide range of possible fitness effects of a single sequence step. The profiles of the pinwheels are similar, indicating the correlation of fitness landscapes, even if WT sequences (centers of the wheels) are only 40% identical and widely separated. Principal component analysis demonstrates a correlation between experimental fitness landscapes and amino-acid preferences in evolved sequences.
Model for the evolution of TIM barrels through gene duplication and domain fusion
, as proposed by Lang
et al
.
[
13
]
This model described the evolution of enzymes HisA and HisF of the histidine biosynthesis pathway. Two gene duplication steps are thought to have occurred. The first gene duplication resulted in two half-barrels that later fused and evolved into an ancestral TIM barrel. The second gene duplication event lead to diversification, and the evolution of different TIM barrel enzymes catalyzing different reactions.
sTIM-11, the first successful
de novo
TIM barrel design.
The asymmetric (αβ)
2
units are colored distinctly, highlighting the internal 4-fold symmetry.