Foraminifera
"Monothalamea" Tubothalamea Globothalamea incertae sedis Foraminifera (/fəˌræməˈnɪfərə/ fə-RAM-ə-NIH-fə-rə; Latin for "hole bearers"; informally called "forams") are single-celled organisms, members of a phylum or class of Rhizarian protists characterized by streaming granular ectoplasm for catching food and other uses; and commonly an external shell (called a "test") of diverse forms and materials.
Most foraminifera are marine, the majority of which live on or within the seafloor sediment (i.e., are benthic, with different sized species playing a role within the macrobenthos, meiobenthos, and microbenthos),[2] while a smaller number float in the water column at various depths (i.e., are planktonic), which belong to the suborder Globigerinina.
[3] Fewer are known from freshwater[4] or brackish[5] conditions, and some very few (nonaquatic) soil species have been identified through molecular analysis of small subunit ribosomal DNA.
[14] The earliest known reference to foraminifera comes from Herodotus, who in the 5th century BCE noted them as making up the rock that forms the Great Pyramid of Giza.
Strabo, in the 1st Century BCE, noted the same foraminifera, and suggested that they were the remains of lentils left by the workers who built the pyramids.
[15] Robert Hooke observed a foraminifera under the microscope, as described and illustrated in his 1665 book Micrographia:I was trying several small and single Magnifying Glasses, and casually viewing a parcel of white Sand, when I perceiv'd one of the grains exactly shap'd and wreath'd like a Shell[...] I view'd it every way with a better Microscope and found it on both sides, and edge-ways, to resemble the Shell of a small Water-Snail with a flat spiral Shell[...][16]Antonie van Leeuwenhoek described and illustrated foraminiferal tests in 1700, describing them as minute cockles; his illustration is recognizable as being Elphidium.
Brady recognized 10 families with 29 subfamilies, with little regard to stratigraphic range; his taxonomy emphasized the idea that multiple different characters must separate taxonomic groups, and as such placed agglutinated and calcareous genera in close relation.
Geologist Irene Crespin undertook extensive research in this field, publishing some ninety papers—including notable work on foraminifera—as sole author as well as more than twenty in collaboration with other scientists.
[25] Prior to the recognition of evolutionary relationships among the members of the Rhizaria, the Foraminifera were generally grouped with other amoeboids as phylum Rhizopodea (or Sarcodina) in the class Granuloreticulosa.
Although as yet unsupported by morphological correlates, molecular data strongly suggest the Foraminifera are closely related to the Cercozoa and Radiolaria, both of which also include amoeboids with complex shells; these three groups make up the Rhizaria.
[33] The foraminiferal cell is divided into granular endoplasm and transparent ectoplasm from which a pseudopodial net may emerge through a single opening or through many perforations in the test.
The pseudopods are used for locomotion, anchoring, excretion, test construction and in capturing food, which consists of small organisms such as diatoms or bacteria.
Forams have evolved special cellular mechanisms to quickly assemble and disassemble microtubules, allowing for the rapid formation and retraction of elongated pseudopodia.
[37] Some species of foraminifera have large, empty vacuoles within their cells; the exact purpose of these is unclear, but they have been suggested to function as a reservoir of nitrate.
[37] Several species of xenophyophore have been found to have unusually high concentrations of radioactive isotopes within their cells, among the highest of any eukaryote.
[38] Modern Foraminifera are primarily marine organisms, but living individuals have been found in brackish, freshwater[34] and even terrestrial habitats.
[52] Nonmarine foraminifera have traditionally been neglected in foram research, but recent studies show them to be substantially more diverse than previously known.
They are known to inhabit disparate ecological niches, including mosses, rivers, lakes and ponds, wetlands, soils, peat bogs, and sand dunes.
The agamont is the asexual reproduction phase of the foraminifera; upon reaching adulthood, the protoplasm entirely vacates the test and divides its cytoplasm meiotically via multiple fission to form a number of haploid offspring.
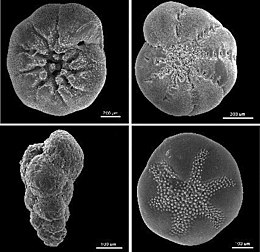
Owing to their generally hard and durable construction (compared to other protists), the tests of foraminifera are a major source of scientific knowledge about the group.
A wide variety of test morphologies is found in both unilocular and multilocular forms, including spiraled, serial, and milioline, among others.
The earliest known definite foraminifera appear in the fossil record towards the very end of the Ediacaran; these forms all have agglutinated tests and are unilocular.
[70][22] Later spirillinids would evolve multilocularity and calcitic tests, with the first such forms appearing during the Triassic; the group saw little effects on diversity due to the K-Pg extinction.
The group remained at low diversity throughout its fossil history; all living representatives belong to the Robertinidae, which first appeared during the Paleocene.
[41][42] Dying planktonic Foraminifera continuously rain down on the sea floor in vast numbers, their mineralized tests preserved as fossils in the accumulating sediment.
[77] The exceptional quality of the fossil record has allowed an impressively detailed picture of species inter-relationships to be developed on the basis of fossils, in many cases subsequently validated independently through molecular genetic studies on extant specimens[78] Because certain types of foraminifera are found only in certain environments, their fossils can be used to figure out the kind of environment under which ancient marine sediments were deposited; conditions such as salinity, depth, oxygenic conditions, and light conditions can be determined from the different habitat preferences of various species of forams.
This allows workers to track changing climates and environmental conditions over time by aggregating information about the foraminifera present.
[86] Although the PETM is now commonly held to be a case study for global warming and massive carbon emission, the cause, details, and overall significance of the event remain uncertain.
[98] For the same reasons they make useful biostratigraphic markers, living foraminiferal assemblages have been used as bioindicators in coastal environments, including indicators of coral reef health.
[99] Agglutinated fossil foraminifera buried deeply in sedimentary basins can be used to estimate thermal maturity, which is a key factor for petroleum generation.


- Test
- Proloculus (first chamber)
- Chambers
- Foramen (older apertures)
- Endoplasm
- Lipid globule
- Nucleus
- Nucleolus
- Endoplasmic reticulum , the transport network for molecules going to specific parts of the cell
- Annular lamellae
- Mitochondrion , creates ATP (energy) for the cell
- Nascent annular lamellae
- Golgi apparatus ; modifies proteins and sends them out of the cell
- Digestive vacuole
- Peroxisome , generates and scavenges hydrogen peroxide
- Phagocytic vacuole
- Lysosome , holds enzymes
- Adhesive substance vesicle
- Aperture (opening of newest chamber)
- Ectoplasm
- Reticulopodia
- Adhesive granules
- Prey