Polyploidy
It may occur due to abnormal cell division, either during mitosis, or more commonly from the failure of chromosomes to separate during meiosis or from the fertilization of an egg by more than one sperm.
[1] In addition, it can be induced in plants and cell cultures by some chemicals: the best known is colchicine, which can result in chromosome doubling, though its use may have other less obvious consequences as well.
Sequencing confirmed that this species originated from E. × robertsii, a sterile triploid hybrid between E. guttata and E. lutea, both of which have been introduced and naturalised in the United Kingdom.
[8] Each of these species experienced independent autopolyploidy events (within-species polyploidy, described below), which then enabled subsequent interspecies gene flow of adaptive alleles, in this case stabilising each young polyploid lineage.
[9] Such polyploidy-enabled adaptive introgression may allow polyploids at act as 'allelic sponges', whereby they accumulate cryptic genomic variation that may be recruited upon encountering later environmental challenges.
[24][25] Rarely, autopolyploids arise from spontaneous, somatic genome doubling, which has been observed in apple (Malus domesticus) bud sports.
[33] This trait is often used as a diagnostic criterion to distinguish autopolyploids from allopolyploids, which commonly display disomic inheritance after they progress past the neopolyploid stage.
[34] While most polyploid species are unambiguously characterized as either autopolyploid or allopolyploid, these categories represent the ends of a spectrum of divergence between parental subgenomes.
Polyploids that fall between these two extremes, which are often referred to as segmental allopolyploids, may display intermediate levels of polysomic inheritance that vary by locus.
If divergence between homoeologous chromosomes is even across the two subgenomes, this can theoretically result in rapid restoration of bivalent pairing and disomic inheritance following allopolyploidization.
However multivalent pairing is common in many recently formed allopolyploids, so it is likely that the majority of meiotic stabilization occurs gradually through selection.
Examples of unexpected but recently confirmed ancient genome duplications include baker's yeast (Saccharomyces cerevisiae), mustard weed/thale cress (Arabidopsis thaliana), rice (Oryza sativa), and two rounds of whole genome duplication (the 2R hypothesis) in an early evolutionary ancestor of the vertebrates (which includes the human lineage) and another near the origin of the teleost fishes.
For example, durum wheat is the result of the inter-species hybridization of two diploid grass species Triticum urartu and Aegilops speltoides.
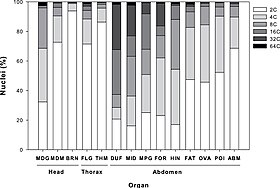
[52] Polyploidy also occurs commonly in amphibians; for example the biomedically important genus Xenopus contains many different species with as many as 12 sets of chromosomes (dodecaploid).
An octodontid rodent of Argentina's harsh desert regions, known as the plains viscacha rat (Tympanoctomys barrerae) has been reported as an exception to this 'rule'.
It was therefore surmised that an Octomys-like ancestor produced tetraploid (i.e., 2n = 4x = 112) offspring that were, by virtue of their doubled chromosomes, reproductively isolated from their parents.
Polyploidy was induced in fish by Har Swarup (1956) using a cold-shock treatment of the eggs close to the time of fertilization, which produced triploid embryos that successfully matured.
[61] John Gurdon (1958) transplanted intact nuclei from somatic cells to produce diploid eggs in the frog, Xenopus (an extension of the work of Briggs and King in 1952) that were able to develop to the tadpole stage.
B. S. Haldane hailed the work for its potential medical applications and, in describing the results, became one of the first to use the word "clone" in reference to animals.
[citation needed] The vast majority of triploid conceptions end as a miscarriage; those that do survive to term typically die shortly after birth.
He failed to exhibit normal mental or physical neonatal development, and died from a Pneumocystis carinii infection, which indicates a weak immune system.
It is unknown whether these embryos fail to implant and are therefore rarely detected in ongoing pregnancies or if there is simply a selective process favoring the diploid cells.
[74] Polyploid plants can arise spontaneously in nature by several mechanisms, including meiotic or mitotic failures, and fusion of unreduced (2n) gametes.
Besides plants and animals, the evolutionary history of various fungal species is dotted by past and recent whole-genome duplication events (see Albertin and Marullo 2012[89] for review).
Several examples of polyploids are known: In addition, polyploidy is frequently associated with hybridization and reticulate evolution that appear to be highly prevalent in several fungal taxa.
As for plants and animals, fungal hybrids and polyploids display structural and functional modifications compared to their progenitors and diploid counterparts.
[101] Phenotypic diversification is also observed following polyploidization and/or hybridization in fungi,[102] producing the fuel for natural selection and subsequent adaptation and speciation.
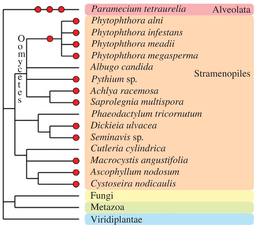
In the Alveolata group, the remarkable species Paramecium tetraurelia underwent three successive rounds of whole-genome duplication[106] and established itself as a major model for paleopolyploid studies.
The mechanism by which the genome is accurately restored involves RecA-mediated homologous recombination and a process referred to as extended synthesis-dependent strand annealing (SDSA).
[110] The archaeon Halobacterium salinarium is polyploid[111] and, like Deinococcus radiodurans, is highly resistant to X-ray irradiation and desiccation, conditions that induce DNA double-strand breaks.