Invasion (cancer)
[2] The results of numerous experimental and clinical studies of malignant neoplasms have indicated that invasive growth and metastasis are the main manifestations of tumor progression, which constitute two closely related processes.
[2] A malignant tumor is defined by its capacity to initiate a biological phenomenon known as the metastatic cascade, a complex multi-stage process in which cell invasion precedes further cancer progression and the formation of metastases in distant organs and tissues.
Due to intratumoral heterogeneity, some tumor cells may undergo regression and death, while others, resilient against opposing microenvironmental factors, acquire an aggressive phenotype and the capacity to metastasize.
This type of migration indicates tissue rearrangement, underlies the processes of embryonic morphogenesis, and is also an essential component of the healing of wound surfaces.
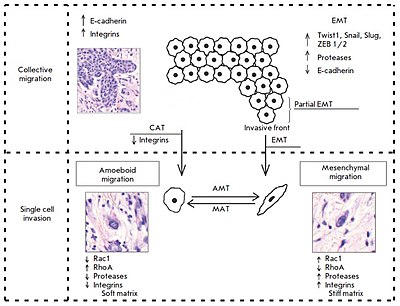
[2] In the case of collective migration, tumor cells form protrusions (pseudopodia) at the leading edge, use integrins to form focal contacts with the actin cytoskeleton, and perform proteolytic degradation of the extracellular matrix, creating a space for invasion of the tumor tissue and extensively involving the actin-myosin contractile apparatus in the process to ensure successful migration.
In in vitro experiments, the introduction of fibroblasts in the culture induces collective tumor cell migration to the underlying matrix in the form of chains.
This protein is known to be involved in the regulation of developing invadopodias, which are structures typical of malignant tumor cells and responsible for the destruction of the surrounding extracellular matrix.
[2] The mesenchymal mechanisms of invasive cell growth, in contrast to the amoeboid type of migration, are characterized by the occurrence of more complex processes and the involvement of a larger number of cellular molecules.
Since malignant cells that use mesenchymal-type movement lose their epithelial polarity and adopt an elongated spindle shape resembling that of fibroblasts, this type of invasion is also referred to as 'fibroblast-like' migration.
This cycle includes the following changes: 1) formation of a protrusion on one of the cell poles – a lamellipodia or a filopodia produced by contractions of the actin cytoskeleton under the control of small GTPases Rac1 and Cdc42 with rapid involvement of integrins of the β1 family; 2) occurrence of focal adhesion with the involvement of integrins β1 and β3 at the contact site between the extracellular matrix and the cell; 3) assembly of focal contacts, which is based on integrin-mediated interactions, and activation of proteolytic enzymes (matrix metalloproteinases, serine and threonine proteases, cathepsins) at the "cell-matrix" interface that leads to the destruction and remodeling of the surrounding extracellular matrix; 4) a change in the actin cytoskeleton polarization under myosin II-mediated control, the occurrence of cell body contractions; and 5) "pulling" the trailing edge toward movement through the newly formed defects in the matrix structure.
The high migration efficiency of a single cell using the mesenchymal mechanism in dense tissues is explained by proteolysis due to the secretion of various proteases and by the ability to form focal contacts with stromal elements.
[2] Based on the suppression of the expression of the relevant genes using small interfering RNAs, the specific activity of GTPases Rac1 and Cdc42 was demonstrated to be the characteristic feature of the mesenchymal type of invasion.
These data most likely indicate that, under conditions of a reduction in or complete loss of their ability to spread to the surrounding tissues using the main molecules that perform adhesion and destruction of the extracellular matrix, tumor cells turn to the amoeboid mechanism of invasion, which becomes the only and most effective mode of migration.
Amoeboid cells are characterized by fast deformability, adaption of their shapes to existing structures of the surrounding extracellular matrix, and penetration through them via narrow spaces in a compressed form.
Developing changes in the cell shape are generated by the cortical actin cytoskeleton that is, in turn, controlled by small GTPase RhoA and its effector, ROCK kinase.
This GTPase belongs to the superfamily of small GTP hydrolases, whose members play key roles in the amoeboid type of invasion, since they are involved in signal transduction and, thereby, in the regulation of a wide variety of processes occurring in the cell, including reorganization of the actin cytoskeleton during migration.
Important aspects are the absence of proteolysis at the sites of cell-matrix interactions and the lack of expression of proteolytic enzymes that destroy the extracellular matrix.
In vitro studies have demonstrated that, in the case of an amoeboid type of invasive growth, it is likely due to these properties that tumor cells are capable of moving at the highest speed in cultures (20 μm/min).
The following mechanisms leading to the transition of cells from the mesenchymal to the amoeboid type of invasive growth (mesenchymal-amoeboid transition) have been described: 1) reduction in or complete abolition of pericellular proteolysis due to application of protease inhibitors; 2) reduction in the activity of integrin receptors and their interactions with surrounding stromal elements by their antagonists; 3) increase in and stabilization of the activity of small GTPase RhoA and its ROCK effector.
[2] The epithelial-mesenchymal transition is a mechanism during which the tumor cell detaches from the epithelial layer and gains motility, the "locomotor phenotype," which promotes invasive growth and metastasis.
During carcinogenesis, epithelial cells are subjected to a morphological transformation that is phenotypically similar to EMT but develops in the absence of the relevant HGF ligand.
[2] Taddei et al. have indicated that EMT develops due to the induction of programs associated with the activation of key transcription factors, such as TWIST1, Snail, Slug, and ZEB1/2.
This results in the disruption of strong cadherin junctions and activation of polar cell migration and proteolysis of extracellular matrix components by various secreted proteases, with the functions of integrin receptors being retained.
[2] Experimental data indicates the potential existence of a collective-amoeboid transition, when tumor masses invading surrounding tissues in the form of collective multicellular groups dissociate into single migrating cells that use the amoeboid movement.
This event has been shown to become possible with the application of inhibitors of integrin receptors of the β1 family, since these molecules play a key role both in the formation of cell-cell contacts and in the interactions between tumor cells and surrounding tissue components.
Nguyen et al. demonstrated that the selective inhibitor PD173074 of the fibroblast growth factor receptor 1 (FGFR1) inhibits the MAPK signaling pathway regulating the activity of the AP-1 protein, which, in turn, induces the development of MET.
For example, there was a decrease in the activity of the genes of cadherins, which are responsible for cell-cell contacts, in the order: solid – alveolar and trabecular structures – discrete groups of tumor cells.
According to research by Krakhmal et al., breast tumors containing both alveolar and trabecular structures, as well as demonstrating significant morphological diversity, are characterized by increased drug resistance.
[2] By the degree of invasion, a cancer can be classified as in situ when malignant cells are present as a tumor but have not metastasized, or invaded beyond the layer or tissue type where it arose.
However, there remain unexplored questions concerning a possible relationship between different types of invasive cell growth and the parameters of lymphogenous and hematogenous metastasis, the features of disease progression, as well as the efficacy of the chosen therapy.