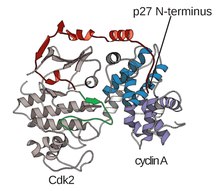
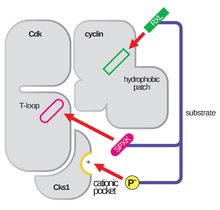
Cyclin-dependent kinase complex
High resolution structures exist for approximately 25 CDK-cyclin complexes in total within the Protein Data Bank.
[4] Based on function, there are two general populations of CDK-cyclin complex structures, open and closed form.
These cyclin binding sites are the regions of highest variability in CDKs despite relatively high sequence homology surrounding the αL-12 Helix motif of this structural component.
This activity is aided by the notable flexibility that the Gly-rich loop has within the structure of most CDK allowing for its rotation toward the activation loop to have a significant effect on reducing substrate affinity without major changes in the overall CDK-cyclin complex structure.
In discussion of the T-loop and the Gly-rich loop, it is important to note that these regions, which must be able to spatially interact in order to carry out their biochemical functions, lie on opposite lobes of the CDK itself.
Given that this region is so conserved across the protein superfamily of kinases, this mechanism where the αC-Helix has been shown to fold out of the N-terminal lobe of the kinase, allowing for increased access to the αL-12 Helix that lies within the T-loop, is considered a potential target for drug development.
From what is known about the complexes formed during each phase of the cell cycle in yeast, proposed models have emerged based on important phosphorylation sites and transcription factors involved.
As previously mentioned, in yeast, only one cyclin-dependent kinase (CDK) is associated with several different cyclins.
[9] Studies have indicated that there is no difference between CDKCs cyclin D1-Cdk4/6, therefore, any unique properties can possibly be linked to substrate specificity or activation.
During late G1 phase, CDKCs bind and phosphorylate members of the retinoblastoma (Rb) protein family.
[11][12] Once phosphorylation occurs, transcription factors are then released to irreversibly inactivate pRB and progression into the S phase of the cell cycle ensues.
[1] Even though the majority of the known CDKCs are involved in the cell cycle, not all kinase complexes function in this manner.
[15][16][17] Additionally, cyclin H-Cdk7 complexes may play a role in meiosis in male germ cells,[18] and has been shown to be involved in transcriptional activities as well.