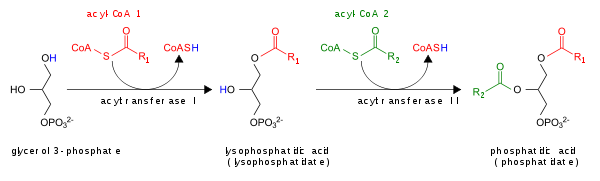
Phosphatidic acid
Phosphatidic acids are anionic phospholipids important to cell signaling and direct activation of lipid-gated ion channels.
However, it is worth noting that in vitro, the various acyltransferases exhibit different substrate specificities with respect to the acyl-CoAs that are incorporated into PA.
This suggests that the various acyltransferases present in mammalian and yeast cells may be responsible for producing different pools of PA.[11] The conversion of PA into diacylglycerol (DAG) by LPPs is the commitment step for the production of phosphatidylcholine (PC), phosphatidylethanolamine (PE) and phosphatidylserine (PS).
In addition, DAG is also converted into CDP-DAG, which is a precursor for phosphatidylglycerol (PG), phosphatidylinositol (PI) and phosphoinositides (PIP, PIP2, PIP3).
PA is a unique phospholipid in that it has a small highly charged head group that is very close to the glycerol backbone.
A major event in the budding of vesicles, such as transport carriers from the Golgi, is the creation and subsequent narrowing of the membrane neck.
Studies have suggested that this process may be lipid-driven, and have postulated a central role for DAG due to its, likewise, unique molecular shape.
LPAAT activity doubles the number of acyl chains, greatly increasing the cross-sectional area of the lipid that lies ‘within’ the membrane while the surface headgroup remains unchanged.
[15] The curvature induced by these lipids was shown to be dependent not only on the structure of lysoPA versus PA but also on dynamic properties, such as the hydration of head groups and inter- and intramolecular interactions.
The neutralisation of the otherwise repulsive charges of the headgroups and the absence of any steric hindrance enables strong intermolecular interactions between the acyl chains, resulting in PA-rich microdomains.
Thus in vitro, physiological changes in pH, temperature, and cation concentrations have strong effects on the membrane curvature induced by PA and lysoPA.
For instance, the Ca2+ requirement for the fusion of complex liposomes is not greatly affected by the addition of annexin I, though it is reduced by PLD.
PA is kept low in the bulk of the membrane in order to transiently burst and signal locally in high concentration.
[16] For example TREK-1 channels are activated by local association with PLD and production of PA.[17] The dissociation constant of PA for TREK-1 is approximately 10 micromolar.
This raises the question as to whether PA has any direct role in signaling or whether it simply acts as a precursor for DAG production.