Non-coding RNA
[18][19][20][21] The cloverleaf secondary structure was finalised following X-ray crystallography analysis performed by two independent research groups in 1974.
[24] Recent notable additions include riboswitches and miRNA; the discovery of the RNAi mechanism associated with the latter earned Craig C. Mello and Andrew Fire the 2006 Nobel Prize in Physiology or Medicine.
The more conserved ncRNAs are thought to be molecular fossils or relics from the last universal common ancestor and the RNA world, and their current roles remain mostly in regulation of information flow from DNA to protein.
The snoRNAs guide covalent modifications of rRNA, tRNA and snRNAs; RNase MRP cleaves the internal transcribed spacer 1 between 18S and 5.8S rRNAs.
Another ubiquitous RNP called SRP recognizes and transports specific nascent proteins to the endoplasmic reticulum in eukaryotes and the plasma membrane in prokaryotes.
In bacteria, Transfer-messenger RNA (tmRNA) is an RNP involved in rescuing stalled ribosomes, tagging incomplete polypeptides and promoting the degradation of aberrant mRNA.
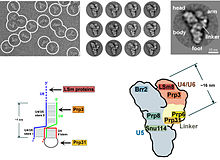
[citation needed] In eukaryotes, the spliceosome performs the splicing reactions essential for removing intron sequences, this process is required for the formation of mature mRNA.
[citation needed] Another group of introns can catalyse their own removal from host transcripts; these are called self-splicing RNAs.
[33] Y RNAs are stem loops, necessary for DNA replication through interactions with chromatin and initiation proteins (including the origin recognition complex).
In the human nucleus, RNase P is required for the normal and efficient transcription of various ncRNAs transcribed by RNA polymerase III.
RNase P exerts its role in transcription through association with Pol III and chromatin of active tRNA and 5S rRNA genes.
[41] A number of ncRNAs are embedded in the 5' UTRs (Untranslated Regions) of protein coding genes and influence their expression in various ways.
[citation needed] RNA leader sequences are found upstream of the first gene of amino acid biosynthetic operons.
These RNA elements form one of two possible structures in regions encoding very short peptide sequences that are rich in the end product amino acid of the operon.
A terminator structure forms when there is an excess of the regulatory amino acid and ribosome movement over the leader transcript is not impeded.
When there is a deficiency of the charged tRNA of the regulatory amino acid the ribosome translating the leader peptide stalls and the antiterminator structure forms.
When iron concentration is low, IRPs bind the ferritin mRNA IRE leading to translation repression.
[citation needed] Internal ribosome entry sites (IRES) are RNA structures that allow for translation initiation in the middle of a mRNA sequence as part of the process of protein synthesis.
[citation needed] Piwi-interacting RNAs (piRNAs) expressed in mammalian testes and somatic cells form RNA-protein complexes with Piwi proteins.
These piRNA complexes (piRCs) have been linked to transcriptional gene silencing of retrotransposons and other genetic elements in germline cells, particularly those in spermatogenesis.
Telomerase is an RNP enzyme that adds specific DNA sequence repeats ("TTAGGG" in vertebrates) to telomeric regions, which are found at the ends of eukaryotic chromosomes.
The enzyme is a reverse transcriptase that carries Telomerase RNA, which is used as a template when it elongates telomeres, which are shortened after each replication cycle.
[66] The miRNAs are involved in the large scale regulation of many protein coding genes,[67][68] the Y RNAs are important for the initiation of DNA replication,[35] telomerase RNA that serves as a primer for telomerase, an RNP that extends telomeric regions at chromosome ends (see telomeres and disease for more information).
Germline mutations in miR-16-1 and miR-15 primary precursors have been shown to be much more frequent in patients with chronic lymphocytic leukemia compared to control populations.
[71] Likewise, a screen of 17 miRNAs that have been predicted to regulate a number of breast cancer associated genes found variations in the microRNAs miR-17 and miR-30c-1of patients; these patients were noncarriers of BRCA1 or BRCA2 mutations, lending the possibility that familial breast cancer may be caused by variation in these miRNAs.
[81] The chromosomal locus containing the small nucleolar RNA SNORD115 gene cluster has been duplicated in approximately 5% of individuals with autistic traits.
[85] Mutations within RNase MRP have been shown to cause cartilage–hair hypoplasia, a disease associated with an array of symptoms such as short stature, sparse hair, skeletal abnormalities and a suppressed immune system that is frequent among Amish and Finnish.
Variation within the seed region of mature miR-96 has been associated with autosomal dominant, progressive hearing loss in humans and mice.
Some publications[24] state that ncRNA and fRNA are nearly synonymous, however others have pointed out that a large proportion of annotated ncRNAs likely have no function.