Restriction point
[1] The defining biochemical feature of the restriction point is the activation of G1/S- and S-phase cyclin-CDK complexes, which in turn phosphorylate proteins that initiate DNA replication, centrosome duplication, and other early cell cycle events.
Originally, Howard Martin Temin showed that chicken cells reach a point at which they are committed to replicate their DNA and are not dependent on extracellular signals.
[4] In 1985, Zetterberg and Larsson discovered that, in all stages of the cell cycle, serum deprivation results in inhibition of protein synthesis.
After the R-point has been passed in G1b, however, extracellular signals are no longer required, and the cell is irreversibly committed to preparing for DNA duplication.
After passing this switch-like “point of no return,” cell cycle completion is no longer dependent on the presence of mitogens.
RTK-associated Grb2 binds Sos, which is a guanine nucleotide exchange factor that converts membrane-bound Ras to its active form (Ras-GDP
[8] Myc also regulates expression of a wide variety of pro-proliferative and pro-growth genes, including some induction of cyclin D2 and Cdk4.
[5] Additionally, sustained ERK activity seems to be important for phosphorylation and nuclear localization of CDK2,[8] further supporting progression through the restriction point.
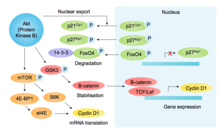
In addition to other pro-growth and pro-survival functions, Akt inhibits glycogen synthase kinase-3β (GSK3β), thereby preventing GSK3β -mediated phosphorylation and subsequent degradation of cyclin D1[10] (see figure[11]).
Extracellular signaling must be maintained, and the cell must also have access to sufficient nutrient supplies to support rapid protein synthesis.
[15] Cyclin D-bound Cdks 4 and 6 are activated by Cdk-activating kinase and drive the cell towards the restriction point.
[14] Similarly, DNA damage and other stressors increase p21 levels, while mitogen-stimulated ERK2 and Akt activity leads to inactivating phosphorylation of p21.
[21] Work by James et al. (2008) further suggests that phosphorylation of tyrosine residues on p27 can switch p27 between an inhibitory and non-inhibitory state while bound to cyclin D-Cdk4/6, offering a model for how p27 is capable of regulating both cyclin-Cdk complex assembly and activity.
A paper published by the Lingchong You and Joe Nevins groups at Duke University in 2008 demonstrated that the a bistable hysteric E2F switch underlies the restriction point.
Serum-starved cells were stimulated with varying serum concentrations, and the GFP readout was recorded at a single-cell level.
By using drugs that inhibit completion of the restriction point, such as growth factor receptor inhibitors, normal cells are prevented from proliferating, and are thus protected from chemotherapy treatments.