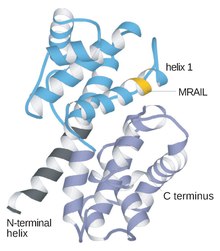
Cyclin
[1] Cyclins were originally discovered by R. Timothy Hunt in 1982 while studying the cell cycle of sea urchins.
Cyclins themselves have no enzymatic activity but have binding sites for some substrates and target the Cdks to specific subcellular locations.
These phosphorylated proteins, in turn, are responsible for specific events during cell division such as microtubule formation and chromatin remodeling.
The Cdk- G1/S cyclin complex begins to induce the initial processes of DNA replication, primarily by arresting systems that prevent S phase Cdk activity in G1.
The cyclins also promote other activities to progress the cell cycle, such as centrosome duplication in vertebrates or spindle pole body in yeast.
The destruction of M cyclins during metaphase and anaphase, after the Spindle Assembly Checkpoint is satisfied, causes the exit of mitosis and cytokinesis.
[8] Kaposi sarcoma herpesvirus (KSHV) encodes a D-type cyclin (ORF72) that binds CDK6 and is likely to contribute to KSHV-related cancers.
[14][15] Notably, recent studies have shown that cyclin A creates a cellular environment that promotes microtubule detachment from kinetochores in prometaphase to ensure efficient error correction and faithful chromosome segregation.
As levels of cyclin A decline, microtubule attachments become stable, allowing the chromosomes to be divided correctly as cell division proceeds.
Leland H. Hartwell, R. Timothy Hunt, and Paul M. Nurse won the 2001 Nobel Prize in Physiology or Medicine for their discovery of cyclin and cyclin-dependent kinase.