G2 phase
Fission yeast (Schizosaccharomyces pombe) has been previously shown to employ such a mechanism, via Cdr2-mediated spatial regulation of Wee1 activity.
[3] Though Wee1 is a fairly conserved negative regulator of mitotic entry, no general mechanism of cell size control in G2 has yet been elucidated.
Biochemically, the end of G2 phase occurs when a threshold level of active cyclin B1/CDK1 complex, also known as Maturation promoting factor (MPF) has been reached.
[5] Mitotic entry is determined by a threshold level of active cyclin-B1/CDK1 complex, also known as cyclin-B1/Cdc2 or the maturation promoting factor (MPF).
Active cyclin-B1/CDK1 triggers irreversible actions in early mitosis, including centrosome separation, nuclear envelope breakdown, and spindle assembly.
[7] Subsequent discussions will focus on the spatial and temporal activation of cyclin B1/CDK in mammalian cells, but similar pathways are applicable in both other metazoans and in S. cerevisiae.
The effects of Wee1 and Myt1 are counteracted by phosphatases in the cdc25 family, which remove the inhibitory phosphates on CDK1 and thus convert the cyclin B1-CDK1 complex to its fully activated form, MPF.
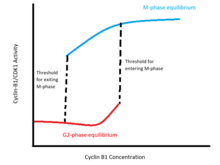
These positive feedback loops encode a hysteretic bistable switch in CDK1 activity relative to cyclin B1 levels (see figure).
Negative feedback provides one essential element of this long-term oscillation: cyclin-B/CDK activates APC/C, which causes degradation of cyclin-B from metaphase onwards, restoring CDK1 to its inactive state.
However, simple negative feedback loops lead to damped oscillations that eventually settle on a steady state.
Recently, evidence has emerged suggesting a more important role for cyclin A2/CDK complexes in regulating the initiation of this switch.
[7] In addition to the bistable and hysteretic aspects of cyclin B1-CDK1 activation, regulation of subcellular protein localization also contributes to the G2/M transition.
In Xenopus laevis, cyclin B1 contains four analogous CRS serine phosphorylation sites (S94, S96, S101, and S113) indicating that this mechanism is highly conserved.
The regulators of these phosphorylation sites are still largely unknown but several factors have been identified, including extracellular signal-regulated kinases (ERKs), PLK1, and CDK1 itself.
It is thought that the simultaneous transport of cdc25 and cyclin-B1/CDK1 into the nucleus amplify the switch-like nature of the transition by increasing the effective concentrations of the proteins.
[7] Experimental approaches to mitigate these changes include both pharmacological inhibition of CDK1 and downregulation of cyclin B1 expression (e.g., via siRNA).
Bypassing the checkpoint leads to the rapid accumulation of deleterious mutations, which is thought to drive the cancerous cells into apoptosis.