SK channel
Their activation limits the firing frequency of action potentials and is important for regulating afterhyperpolarization in the neurons of the central nervous system as well as many other types of electrically excitable cells.
This is accomplished through the hyperpolarizing leak of positively charged potassium ions along their concentration gradient into the extracellular space.
[3] SK channels are thought to be involved in synaptic plasticity and therefore play important roles in learning and memory.
[12] Additional association of the phosphorylating kinase CK2 and dephosphorylating phosphatase PP2A on the cytoplasmic face of the protein allow for enriched Ca2+-sensitivity—and thus—kinetics modulation.
[13] CK2 serves to phosphorylate the SKCa-bound CaM at the T80 residue, rather than the channel helices themselves, to reduce calcium sensitivity.
Calmodulin pushes the S4-S5 linker to allow the expansion of the S6 bundle crossing, leading to opening of the pore.
[17] In addition, SK channels (SK1-SK3) but not SK4 (IK) are sensitive to blockade by the bee toxin apamin,[18] and the scorpion venoms tamapin and charybdotoxin (ChTx), all via competitive antagonism for access to the mouth of the pore formation.
Examples include: In dendritic spines, SK channels are directly coupled to NMDA receptors.
[12] Experiments using apamin have shown that specifically blocking SK channels can increase learning and long-term potentiation.
In addition, brain-derived neurotrophic factor (BDNF) causes the down-regulation of SK channels, which facilitates long-term potentiation.
[5] An increase in SK channel activity that occurs over time may be related to decreases in plasticity and memory that is seen with aging.
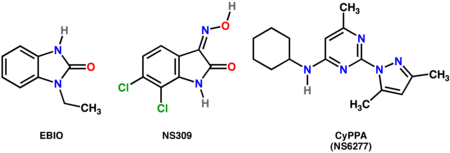
Multiple pharmacological techniques have been used to adjust SK affinity for calcium ions, thereby modulating the excitability of substantia nigra dopaminergic neurons.
In addition, techniques have been used to modulate SK channels in order to alter the dopamine phenotype of neurons.