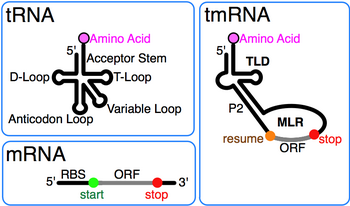
Transfer-messenger RNA
The tmRNA is remarkably versatile: it recycles the stalled ribosome, adds a proteolysis-inducing tag to the unfinished polypeptide, and facilitates the degradation of the aberrant messenger RNA.
In other bacterial species, a permuted ssrA gene produces a two-piece tmRNA in which two separate RNA chains are joined by base-pairing.
[3] Subsequent sequence comparison revealed the full tRNA-like domain (TLD) formed by the 5' and 3' ends of tmRNA, including the acceptor stem with elements like those in alanine tRNA that promote its aminoacylation by alanine-tRNA ligase.
[5][6] Watson-Crick and G-U base pairs were identified by comparing the bacterial tmRNA sequences using automated computational methods in combination with manual alignment procedures.
[7][10] Circularly permuted ssrA has been reported in three major lineages: i) all alphaproteobacteria and the primitive mitochondria of jakobid protists, ii) two disjoint groups of cyanobacteria (Gloeobacter and a clade containing Prochlorococcus and many Synechococcus), and iii) some members of the betaproteobacteria (Cupriavidus and some Rhodocyclales).
In mitochondria, the MLR has been lost, and a remarkable re-permutation of mitochondrial ssrA results in a small one-piece product in Jakoba libera.
High-resolution structures of the complete tmRNA molecules are currently unavailable and may be difficult to obtain due to the inherent flexibility of the MLR.
[19] Coding by tmRNA was discovered in 1995[20] when Simpson and coworkers overexpressed the mouse cytokine IL-6 in E. coli and found multiple truncated cytokine-derived peptides each tagged at the carboxyl termini with the same 11-amino acid residue extension (A)ANDENYALAA.
[21] While details of the trans-translation mechanism are under investigation it is generally agreed that tmRNA first occupies the empty A site of the stalled ribosome.
Trans-translation is essential in some bacterial species, whereas other bacteria require tmRNA to survive when subjected to stressful growth conditions.
By different strategies none of these disrupt gene function: group I introns remove themselves by self-splicing, rickettsial palindromic elements (RPEs) insert in innocuous sites, and integrase-encoding genomic islands split their target ssrA yet restore the split-off portion.
[11] Subsequently, the presence of a mitochondrial gene (ssrA) coding for tmRNA, as well as transcription and RNA processing sites were confirmed for all but one member of jakobids.
[30] Like in α-Proteobacteria (the ancestors of mitochondria), mt-tmRNAs are circularly permuted, two-piece RNA molecules, except in Jakoba libera where the gene has reverted to encoding a one-piece tmRNA conformation.
[13] Mitochondrial tmRNA genes were initially recognized as short sequences that are conserved among jakobids and that have the potential to fold into a distinct tRNA-like secondary structure.
Finally, instead of the tRNA-like D-stem with a shortened three-nucleotide D-loop characteristic for bacterial tmRNAs, mitochondrial counterparts have a highly variable 5 to 14-nt long loop.
RNA-Seq data of Phytophthora sojae show an expression level similar to that of neighboring mitochondrial tRNAs, and four major processing sites confirm the predicted termini of mature mt-tmRNA.