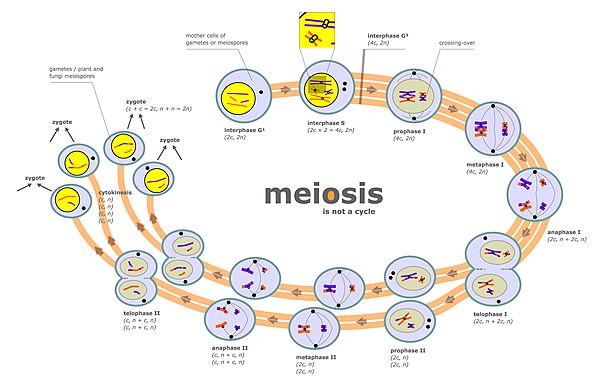
Chromosome segregation
These chromatids separate to opposite poles, a process facilitated by a protein complex referred to as cohesin.
During this stage, segregation occurs by a process similar to that during mitosis, except that in this case prophase II is not preceded by a round of DNA replication.
This is because, at the end of meiotic prophase I, CO recombination provides a physical link that holds homologous chromosome pairs together.
Together with cohesion linkage between sister chromatids, CO recombination may help ensure the orderly segregation of the paired homologous chromosomes to opposite poles.
Mutants of S. cerevisiae defective in CO recombination at the level of Holliday junction resolution were found to efficiently undergo proper chromosome segregation.
The pathway that produces the majority of COs in S. cerevisiae, and possibly in mammals, involves a complex of proteins including the MLH1-MLH3 heterodimer (called MutL gamma).
[8] It is an endonuclease that makes single-strand breaks in supercoiled double-stranded DNA,[8][9] and promotes the formation of CO recombinants.
[11] The MSH4/MSH5 complex binds and stabilizes double Holliday junctions and promotes their resolution into crossover products.
[14] Nevertheless, this mutant gave rise to spore viability patterns suggesting that segregation of non-exchange chromosomes occurred efficiently.
[15] This ability depends on the microtubule motor dynein that regulates the movement of chromosomes to the poles of the meiotic spindle.