DREAM complex
[3][4] Studies in Drosophila melanogaster ovarian follicle cells identified a protein complex that bound to repeatedly amplifying chorion genes.
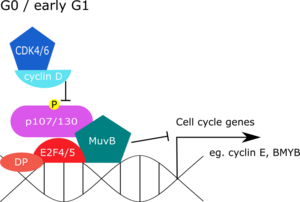
[5] Further studies in the fly embryo nuclear extracts confirmed the coexistence of these proteins with others such as the RB homologues Rbf1 and Rbf2, and others like E2f and Dp.
[7] A protein complex similar to dREAM was subsequently identified in C. elegans extract containing DP, RB, and MuvB, and was named as DRM.
Subsequent immunoprecipitation with BMYB yielded all the MuvB core proteins, but not other members of the DREAM complex – p130, p107, E2F4/5 and DP.
Entry into the cell cycle dissociates p130 from the complex and leads to subsequent recruitment of activating E2F proteins.
In mammalian systems, chromatin-immunoprecipitation (ChIP) studies have revealed that DREAM components are found together at promoters of genes that peak in G1/S or G2/M phase.
[9] Abrogation of the DREAM complex on the other hand, led to increased expression of E2F regulated genes normally repressed in the G0 phase.
Compacted DNA at transcription start sites inhibit gene expression by blocking the docking of RNA polymerase.
[22] Like its counterpart, RB-E2F, the DREAM complex is also affected by similar growth stimuli and subsequent cyclin-CDK activity.
[29][30] Inhibition of PP2a activity reduced promoter binding of some of the proteins of the DREAM complex in the subsequent G1 phase and de-repression of gene expression.
Substitution of this serine to alanine led to reduced binding of the MuvB core to p130 and impaired the ability of cells to enter quiescence.
[33] Due to its regulatory role in the cell cycle, targeting the DREAM complex might enhance anticancer treatments such as imatinib.