Fatty acid synthesis
[2][3] The diagrams presented show how fatty acids are synthesized in microorganisms and list the enzymes found in Escherichia coli.
[2] These reactions are performed by fatty acid synthase II (FASII), which in general contain multiple enzymes that act as one complex.
FASII is less efficient than FASI; however, it allows for the formation of more molecules, including "medium-chain" fatty acids via early chain termination.
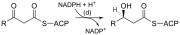
[4] Once formed, the 16:0 carbon fatty acid can undergo a number of modifications, resulting in desaturation and/or elongation.
NADPH is also formed by the pentose phosphate pathway which converts glucose into ribose, which can be used in synthesis of nucleotides and nucleic acids, or it can be catabolized to pyruvate.
[7] In humans, fatty acids are formed from carbohydrates predominantly in the liver and adipose tissue, as well as in the mammary glands during lactation.
The pyruvate produced by glycolysis is an important intermediary in the conversion of carbohydrates into fatty acids and cholesterol.
A young adult human's fat stores average between about 15–20 kg (33–44 lb), but varies greatly depending on age, sex, and individual disposition.
Malate is the only substance that can be removed from the mitochondrion to enter the gluconeogenic pathway to form glucose or glycogen in the liver or any other tissue.
[13] High plasma levels of insulin in the blood plasma (e.g. after meals) cause the dephosphorylation of acetyl-CoA carboxylase, thus promoting the formation of malonyl-CoA from acetyl-CoA, and consequently the conversion of carbohydrates into fatty acids, while epinephrine and glucagon (released into the blood during starvation and exercise) cause the phosphorylation of this enzyme, inhibiting lipogenesis in favor of fatty acid oxidation via beta-oxidation.
[16] Clostridia are the main exception; they have a novel enzyme, yet to be identified, that catalyzes the formation of the cis double bond.
[8] In mammals the aerobic desaturation is catalyzed by a complex of three membrane-bound enzymes (NADH-cytochrome b5 reductase, cytochrome b5, and a desaturase).
These enzymes allow molecular oxygen, O2, to interact with the saturated fatty acyl-CoA chain, forming a double bond and two molecules of water, H2O.
[7] These mammalian enzymes are, however, incapable of introducing double bonds at carbon atoms beyond C-9 in the fatty acid chain.
(Arachidonic acid is the precursor of prostaglandins which fulfill a wide variety of functions as local hormones.
Two DesR-P proteins will dimerize and bind to the DNA promoters of the des gene and recruit RNA polymerase to begin transcription.
Another pathway uses two proteins, DesC and DesB, together to act as a Δ9-desaturase, which inserts a double bond into a saturated fatty acid-CoA molecule.
DesT is also a repressor of fabAB expression for anaerobic desaturation when in presence of exogenous unsaturated fatty acids.
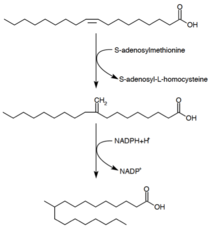
[21][23] Branched chain fatty acids are usually saturated and are found in two distinct families: the iso-series and anteiso-series.
This system is distinct from the branched-chain fatty acid synthetase that utilizes short-chain acyl-CoA esters as primers.
[26] Decarboxylation of the primer precursors occurs through the branched-chain α-keto acid decarboxylase (BCKA) enzyme.
[27] The major end products are 12–17 carbon branched-chain fatty acids and their composition tends to be uniform and characteristic for many bacterial species.
The proportions of these branched-chain fatty acids tend to be uniform and consistent among a particular bacterial species but may be altered due to changes in malonyl-CoA concentration, temperature, or heat-stable factors (HSF) present.
In general, this method is used by bacteria that do not have the ability to perform the branch-chain fatty acid system using alpha-keto primers.
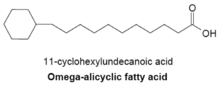
In bacteria with membranes composed mainly of omega-alicyclic fatty acids, the supply of cyclic carboxylic acid-CoA esters is much greater than that of branched-chain primers.
[31] A role as a mediator in intracellular signal transduction is also assumed, as the levels of bioactive lipids, such as lysophospholipids and sphingolipids, correlate with mtFASII.
[1] Disorders in mtFASII lead to the following metabolic diseases: The positions of the double bonds in a fatty acid chain can, therefore, be indicated in two ways, using the C-n or the ω-n notation.
Omega (ω) is the last letter in the Greek alphabet, and is therefore used to indicate the "last" carbon atom in the fatty acid chain.