Magnesium transporter
All forms of life require magnesium, yet the molecular mechanisms of Mg2+ uptake from the environment and the distribution of this vital element within the organism are only slowly being elucidated.
In 1968, Lusk[20] described the limitation of bacterial (Escherichia coli) growth on Mg2+-poor media, suggesting that bacteria required Mg2+ and were likely to actively take this ion from the environment.
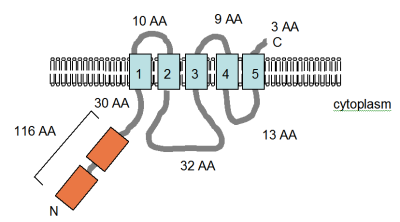
With the large number of complete genome sequences now available for prokaryotes, CorA has been shown to be virtually ubiquitous among the Eubacteria, as well as being widely distributed among the Archaea.
[28] In the more distantly related genes, the similarity is often restricted to the C-terminal part of the protein, and a short amino acid motif GMN within this region is very highly conserved.
A strain of S. typhimurium containing a functional CorA gene but lacking MgtA and MgtB was constructed[37](also see below), and the uptake kinetics of the transporter were analysed.
The km values for Co2+ and Ni2+ are significantly above those expected to be encountered by the cells in their normal environment, so it is unlikely that the CorA transport system mediates the uptake of these ions under natural conditions.
Recently the Mg2+-dependent fluorescence of mag-fura 2 was used to measure the free Mg2+ content of S. typhimurium cells in response to external Mg2+, which showed that CorA is the major uptake system for Mg2+ in bacteria.
[18] The efflux of Mg2+ from bacterial cells was first observed by Lusk and Kennedy (1969)[21] and is mediated by the CorA Mg2+ transport system in the presence of high extracellular concentrations of Mg2+.
[41] The additional strength of the Co(III)Hex inhibition might come from the blocking of the transport pore due to the inability of the protein to ‘dehydrate’ the substrate.
Although the proteins have km values similar to CorA and transport rates approximately 10 times less, the genes may be part of a Mg2+ scavenging system.
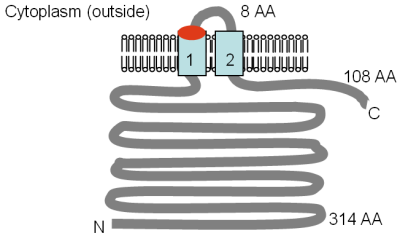
[51] The figure (right), adapted from Smith et al. (1995)[4] and the PFAM database entry, shows the computer-predicted membrane topology of the MgtE protein in Bacillus firmus OF4.
[53] The MgtE gene was first identified by Smith et al. (1995) during a screen for CorA-like proteins in bacteria and complements the Mg2+-uptake-deficient S. typhimurium strain MM281 (corA mgtA mgtB), restoring wild type growth on standard media.
[60] 57Co2+ is taken up into yeast via the Alr1p protein with a km of 77 – 105 μM (;[56] C. MacDiarmid and R. C. Gardner, unpublished data), but the Ki for Mg2+ inhibition of this transport is currently unknown.
Additionally, two electrophysiological studies where Alr1p was produced in yeast or Xenopus oocytes showed a Mg2+-dependent current in the presence of the protein;[62] Salih et al., in prep.
Consistent with this interpretation, the Mnr2 protein was localized to the membrane of the vacuole, an internal compartment implicated in the storage of excess mineral nutrients by yeast.
A direct role of Mnr2 in Mg2+ transport was suggested by the observation that increased Mnr2 expression, which redirected some Mnr2 protein to the cell surface, also suppressed the Mg2+-requirement of an alr1 alr2 double mutant strain.
[6] Gregan et al. (2001a) identified LPE10 by homology to MRS2 and showed that both LPE10 and MRS2 mutants altered the Mg2+ content of yeast mitochondria and affected RNA splicing activity in the organelle.
[7] The open reading frame for the XNTA gene is 1707 bp in size, contains two introns and produces a predicted protein of 550 amino acids.
Only in the last 25 years has a series of reports begun to challenge this view, with new methodologies finding that free Mg2+ content is maintained at levels where changes might influence cellular metabolism.
Very little is known about the Mg2+ transport characteristics of the protein in mammals, but Zsurka et al. (2001) has shown that the human Mrs2p complements the mrs2 mutants in the yeast mitochondrial Mg2+ uptake system.
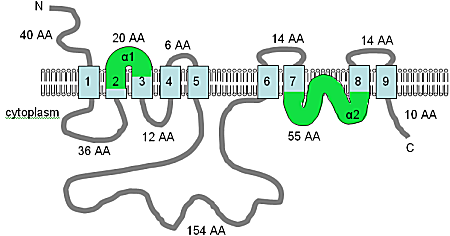
[8] The identification of this gene family in the metazoa began with a signal sequence trap method for isolating secreted and membrane proteins.
The TM domains are shown in light blue, the orientation in the membrane and the positions of the N- and C-termini are indicated, and the figure is not drawn to scale.
The conclusions of the Voets et al. (2004)[83] paper are probably incorrect in attributing the Mg2+ dependent currents to TRPM7 alone, and their kinetic data are likely to reflect the combined TRPM7/ TRPM6 channel.
The report presents a robust collection of data consistent with a channel-like activity passing Mg2+, based on both electrophysiological techniques and also mag-fura 2 to determine changes in cytoplasmic free Mg2+.
Independently, Li et al. (2001)[14] published a report identifying the family and showing that two additional members could complement Mg2+ transport deficient mutants, one in S. typhimurium and the other in S. cerevisiae.
The AtMRS2-1 gene, when expressed in yeast from the MRS2 promoter and being fused C-terminally to the first 95 amino acids of the Mrs2p protein, was directed to the mitochondria, where it complemented a Δmrs2 mutant both phenotypically (mitochondrial RNA splicing was restored) and with respect to the Mg2+ content of the organelle.
[14] An electrophysiological (voltage clamp) analysis of the AtMRS2-11 protein in Xenopus oocytes also showed a Mg2+-dependent current at membrane potentials (ΔΨ) of –100 – –150 mV inside.
The most likely reason for the lack of a definitive localisation of AtMRS2-1 in the Schock et al. paper is that the authors removed the TM domains from the protein, thereby precluding its insertion into a membrane.
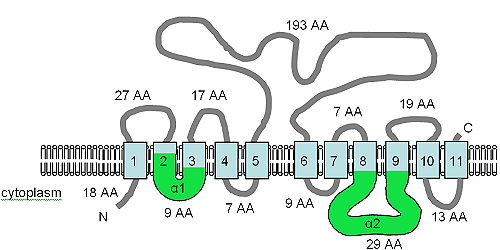
AtMHX is quite closely related to the SLC8 family at the amino acid level and shares a topology with eleven predicted TM domains (Figure A10.5).
The TM domains are shown in light blue, the orientation in the membrane and the positions of the N- and C-termini are indicated, and the figure is not drawn to scale.