Glycolysis
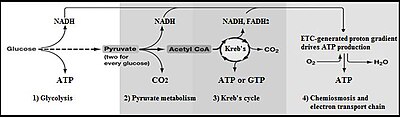
The free energy released in this process is used to form the high-energy molecules adenosine triphosphate (ATP) and reduced nicotinamide adenine dinucleotide (NADH).
[3] The most common type of glycolysis is the Embden–Meyerhof–Parnas (EMP) pathway, which was discovered by Gustav Embden, Otto Meyerhof, and Jakub Karol Parnas.
[citation needed] In high-oxygen (aerobic) conditions, eukaryotic cells can continue from glycolysis to metabolise the pyruvate through the citric acid cycle or the electron transport chain to produce significantly more ATP.
[9] His experiments showed that alcohol fermentation occurs by the action of living microorganisms, yeasts, and that glucose consumption decreased under aerobic conditions (the Pasteur effect).
Harden and Young deduced that this process produced organic phosphate esters, and further experiments allowed them to extract fructose diphosphate (F-1,6-DP).
Meyerhof and his team were able to extract different glycolytic enzymes from muscle tissue, and combine them to artificially create the pathway from glycogen to lactic acid.
Meyerhoff and Junowicz found that the equilibrium constant for the isomerase and aldoses reaction were not affected by inorganic phosphates or any other cozymase or oxidizing enzymes.
[17] The biggest difficulties in determining the intricacies of the pathway were due to the very short lifetime and low steady-state concentrations of the intermediates of the fast glycolytic reactions.
GlucoseHexokinaseGlucose 6-phosphateGlucose-6-phosphateisomeraseFructose 6-phosphatePhosphofructokinase-1Fructose 1,6-bisphosphateFructose-bisphosphatealdolaseDihydroxyacetone phosphate+Glyceraldehyde 3-phosphateTriosephosphateisomerase2 × Glyceraldehyde 3-phosphateGlyceraldehyde-3-phosphatedehydrogenase2 × 1,3-BisphosphoglyceratePhosphoglycerate kinase2 × 3-PhosphoglyceratePhosphoglycerate mutase2 × 2-PhosphoglyceratePhosphopyruvatehydratase (enolase)2 × PhosphoenolpyruvatePyruvate kinase2 × PyruvateThe first five steps of Glycolysis are regarded as the preparatory (or investment) phase, since they consume energy to convert the glucose into two three-carbon sugar phosphates[5] (G3P).
In animals, an isozyme of hexokinase called glucokinase is also used in the liver, which has a much lower affinity for glucose (Km in the vicinity of normal glycemia), and differs in regulatory properties.
The different substrate affinity and alternate regulation of this enzyme are a reflection of the role of the liver in maintaining blood sugar levels.
Because the reaction catalyzed by phosphofructokinase 1 (PFK-1) is coupled to the hydrolysis of ATP (an energetically favorable step) it is, in essence, irreversible, and a different pathway must be used to do the reverse conversion during gluconeogenesis.
Furthermore, the second phosphorylation event is necessary to allow the formation of two charged groups (rather than only one) in the subsequent step of glycolysis, ensuring the prevention of free diffusion of substrates out of the cell.
[19] Cofactors: Mg2+ Destabilizing the molecule in the previous reaction allows the hexose ring to be split by aldolase into two triose sugars: dihydroxyacetone phosphate (a ketose), and glyceraldehyde 3-phosphate (an aldose).
[21] Conversely, triglycerides can be broken down into fatty acids and glycerol; the latter, in turn, can be converted into dihydroxyacetone phosphate, which can enter glycolysis after the second control point.
[33] The phosphorylation and dephosphorylation of these enzymes (ultimately in response to the glucose level in the blood) is the dominant manner by which these pathways are controlled in the liver, fat, and muscle cells.
The liver is also capable of releasing glucose into the blood between meals, during fasting, and exercise thus preventing hypoglycemia by means of glycogenolysis and gluconeogenesis.
In addition hexokinase and glucokinase act independently of the hormonal effects as controls at the entry points of glucose into the cells of different tissues.
[33] Phosphofructokinase is an important control point in the glycolytic pathway, since it is one of the irreversible steps and has key allosteric effectors, AMP and fructose 2,6-bisphosphate (F2,6BP).
However, it is doubtful that this is a meaningful effect in vivo, because citrate in the cytosol is utilized mainly for conversion to acetyl-CoA for fatty acid and cholesterol synthesis.
This process also occurs in animals under hypoxic (or partially anaerobic) conditions, found, for example, in overworked muscles that are starved of oxygen.
Anoxic regeneration of NAD+ is only an effective means of energy production during short, intense exercise in vertebrates, for a period ranging from 10 seconds to 2 minutes during a maximal effort in humans.
(At lower exercise intensities it can sustain muscle activity in diving animals, such as seals, whales and other aquatic vertebrates, for very much longer periods of time.)
The pH in the cytoplasm quickly drops when hydrogen ions accumulate in the muscle, eventually inhibiting the enzymes involved in glycolysis.
In aerobic eukaryotes, a complex mechanism has developed to use the oxygen in air as the final electron acceptor, in a process called oxidative phosphorylation.
[47] Cholesterol can be used as is, as a structural component of cellular membranes, or it can be used to synthesize the steroid hormones, bile salts, and vitamin D.[35][46][47] Pyruvate molecules produced by glycolysis are actively transported across the inner mitochondrial membrane, and into the matrix where they can either be oxidized and combined with coenzyme A to form CO2, acetyl-CoA, and NADH,[35] or they can be carboxylated (by pyruvate carboxylase) to form oxaloacetate.
Hence the addition of oxaloacetate greatly increases the amounts of all the citric acid intermediates, thereby increasing the cycle's capacity to metabolize acetyl CoA, converting its acetate component into CO2 and water, with the release of enough energy to form 11 ATP and 1 GTP molecule for each additional molecule of acetyl CoA that combines with oxaloacetate in the cycle.
NADPH is also formed by the pentose phosphate pathway which converts glucose into ribose, which can be used in synthesis of nucleotides and nucleic acids, or it can be catabolized to pyruvate.
However, some mutations (glycogen storage diseases and other inborn errors of carbohydrate metabolism) are seen with one notable example being pyruvate kinase deficiency, leading to chronic hemolytic anemia.
One such theory suggests that the increased glycolysis is a normal protective process of the body and that malignant change could be primarily caused by energy metabolism.