Scaffold/matrix attachment region
An increased propensity to separate the DNA strands (the so-called 'stress induced duplex destabilization' potential, SIDD) can serve the formation of secondary structures such as cruciforms or slippage structures, which are recognizable features for a number of enzymes (DNAses, topoisomerases, poly(ADP-ribosyl) polymerases and enzymes of the histone-acetylation and DNA-methylation apparatus).
While the number of S/MARs in the human genome has been estimated to approach 64,000 (chromatin domains) plus an additional 10,000 (replication foci), in 2007 still only a minor fraction (559 for all eukaryotes) had met the standard criteria for an annotation in the S/MARt database.
[1] Current views of the nuclear matrix envision it as a dynamic entity, which changes its properties along the requirements of the cell nucleus—much the same as the cytoskeleton adapts its structure and function to external signals.
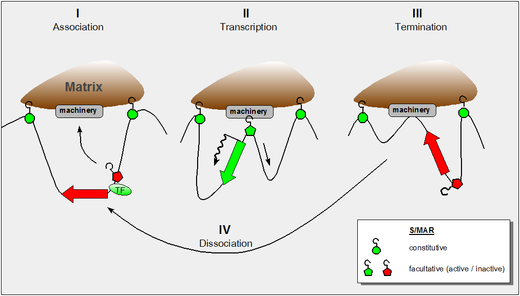
In retrospect it is of note that the discovery of S/MARs has two major routes: Subsequent work demonstrated both the constitutive (SAR-like) and the facultative (MAR-like) function of the elements depending on the context.
The figure shows our present understanding of these properties and it incorporates the following findings: As an alternative to viral vectors, which can have unwanted effects in patients body, non-viral methods of gene therapy are being studied.