Color vision
Color perception is a part of the larger visual system and is mediated by a complex process between neurons that begins with differential stimulation of different types of photoreceptors by light entering the eye.
Those photoreceptors then emit outputs that are propagated through many layers of neurons ultimately leading to higher cognitive functions in the brain.
Each individual cone contains pigments composed of opsin apoprotein covalently linked to a light-absorbing prosthetic group: either 11-cis-hydroretinal or, more rarely, 11-cis-dehydroretinal.
Rather, the perception of color is achieved by a complex process that starts with the differential output of these cells in the retina and which is finalized in the visual cortex and associative areas of the brain.
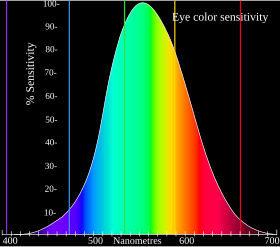
For example, while the L cones have been referred to simply as red receptors, microspectrophotometry has shown that their peak sensitivity is in the greenish-yellow region of the spectrum.
[11] It states that the visual system interprets color in an antagonistic way: red vs. green, blue vs. yellow, black vs. white.
[13] According to Land's Retinex theory, color in a natural scene depends upon the three sets of cone cells ("red," "green," and "blue") separately perceiving each surface's relative lightness in the scene and, together with the visual cortex, assigning color based on comparing the lightness values perceived by each set of cone cells.
X chromosome inactivation means that while only one opsin is expressed in each cone cell, both types may occur overall, and some women may therefore show a degree of tetrachromatic color vision.
[20] Variations in OPN1MW, which encodes the opsin expressed in M cones, appear to be rare, and the observed variants have no effect on spectral sensitivity.
Some midget retinal ganglion cells oppose L and M cone activity, which corresponds loosely to red–green opponency, but actually runs along an axis from blue-green to magenta.
M- and P-cells receive relatively balanced input from both L- and M-cones throughout most of the retina, although this seems to not be the case at the fovea, with midget cells synapsing in the P-laminae.
Many cells in V1 respond to some parts of the spectrum better than others, but this "color tuning" is often different depending on the adaptation state of the visual system.
Such "double-opponent" cells were initially described in the goldfish retina by Nigel Daw;[23][24] their existence in primates was suggested by David H. Hubel and Torsten Wiesel, first demonstrated by C.R.
[26] As Margaret Livingstone and David Hubel showed, double opponent cells are clustered within localized regions of V1 called blobs, and are thought to come in two flavors, red–green and blue-yellow.
Modeling studies have shown that double-opponent cells are ideal candidates for the neural machinery of color constancy explained by Edwin H. Land in his retinex theory.
Although most people are assumed to have the same mapping, the philosopher John Locke recognized that alternatives are possible, and described one such hypothetical case with the "inverted spectrum" thought experiment.
Synesthesia (or ideasthesia) provides some atypical but illuminating examples of subjective color experience triggered by input that is not even light, such as sounds or shapes.
[39] Psychophysical experiments have shown that color is perceived before the orientation of lines and directional motion by as much as 40ms and 80 ms respectively, thus leading to a perceptual asynchrony that is demonstrable with brief presentation times.
In color science, chromatic adaptation is the estimation of the representation of an object under a different light source from the one in which it was recorded.
[50] In view of the importance of color vision to bees one might expect these receptor sensitivities to reflect their specific visual ecology; for example the types of flowers that they visit.
However, the main groups of hymenopteran insects excluding ants (i.e., bees, wasps and sawflies) mostly have three types of photoreceptor, with spectral sensitivities similar to the honeybee's.
[52] The most complex color vision system in the animal kingdom has been found in stomatopods (such as the mantis shrimp) having between 12 and 16 spectral receptor types thought to work as multiple dichromatic units.
There is some evidence that a few mammals, such as cats, have redeveloped the ability to distinguish longer wavelength colors, in at least a limited way, via one-amino-acid mutations in opsin genes.
[59] The adaptation to see reds is particularly important for primate mammals, since it leads to the identification of fruits, and also newly sprouting reddish leaves, which are particularly nutritious.
[citation needed] Color perception mechanisms are highly dependent on evolutionary factors, of which the most prominent is thought to be satisfactory recognition of food sources.
On the other hand, nocturnal mammals have less-developed color vision since adequate light is needed for cones to function properly.
The evolution of trichromatic color vision in primates occurred as the ancestors of modern monkeys, apes, and humans switched to diurnal (daytime) activity and began consuming fruits and leaves from flowering plants.
It allows birds to spot small prey from a distance, navigate, avoid predators, and forage while flying at high speeds.
Finally, since a beam of light can be composed of many different wavelengths, to determine the extent to which a physical color C in Hcolor stimulates each cone cell, we must calculate the integral (with respect to w), over the interval [Wmin,Wmax], of C(w)·s(w), of C(w)·m(w), and of C(w)·l(w).
By considering all the resulting combinations of intensities (IS, IM, IL) as a subset of 3-space, a model for human perceptual color space is formed.