MHC class I
The MHC I: peptide complex is then inserted via the endoplasmic reticulum into the external plasma membrane of the cell.
Alternatively, class I MHC itself can serve as an inhibitory ligand for natural killer cells (NKs).
Reduction in the normal levels of surface class I MHC, a mechanism employed by some viruses[4] and certain tumors to evade CTL responses, activates NK cell killing.
[5] PirB is expressed in the central nervous system and diminishes ocular dominance plasticity in the developmental critical period and adulthood.
[5] PirB loss of function mutant mice also exhibited enhanced plasticity after monocular deprivation during the critical period.
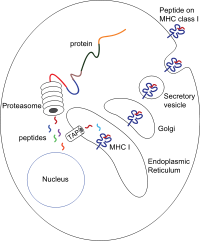
They have membrane proximal Ig fold The peptide translocation from the cytosol into the lumen of the ER is accomplished by the transporter associated with antigen processing (TAP).
TAP binds peptides on the cytoplasmic side and translocates them under ATP consumption into the lumen of the ER.
The MHC molecule lacking a bound peptide is inherently unstable and requires the binding of the chaperones calreticulin and Erp57.
Once the peptide is loaded onto the MHC class I molecule, the complex dissociates and it leaves the ER through the secretory pathway to reach the cell surface.
[18] MHC class I molecules are loaded with peptides generated from the degradation of ubiquitinated cytosolic proteins in proteasomes.
As viruses induce cellular expression of viral proteins, some of these products are tagged for degradation, with the resulting peptide fragments entering the endoplasmic reticulum and binding to MHC I molecules.
As an evolutionary response to this method of immune surveillance, many viruses are able to down-regulate or otherwise prevent the presentation of MHC class I molecules on the cell surface.
[2] Since their emergence in jawed vertebrates, this gene family has been subjected to many divergent evolutionary paths as speciation events have taken place.